

Progressive Differentiation and Instructive Capacities of Amniotic Fluid and Cerebrospinal Fluid Proteomes following Neural Tube Closure
- PMID: 26702835
- PMCID: PMC4691285
- DOI: 10.1016/j.devcel.2015.11.015
Progressive Differentiation and Instructive Capacities of Amniotic Fluid and Cerebrospinal Fluid Proteomes following Neural Tube Closure
Abstract
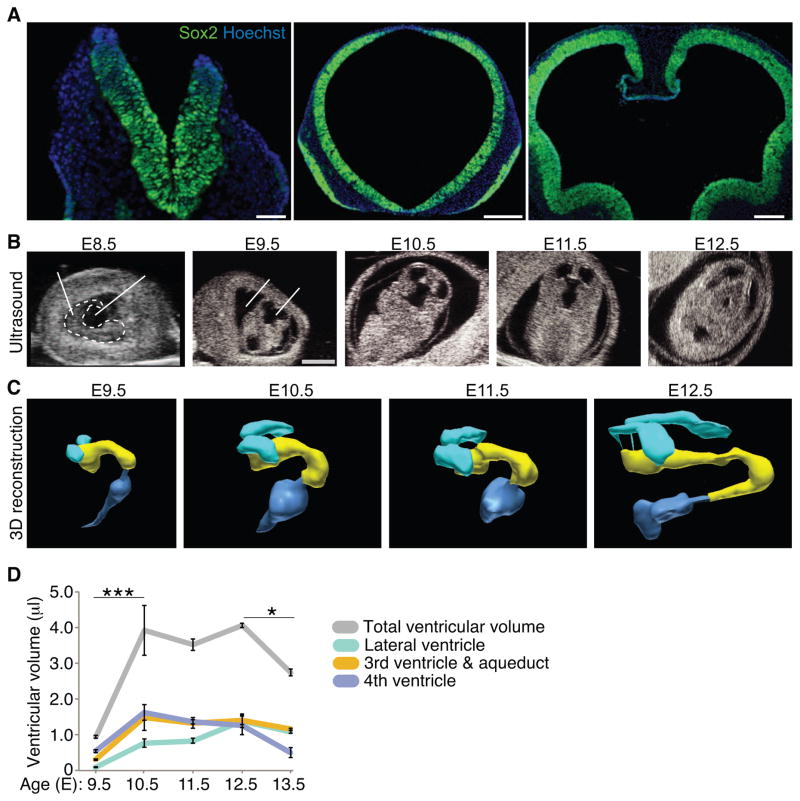
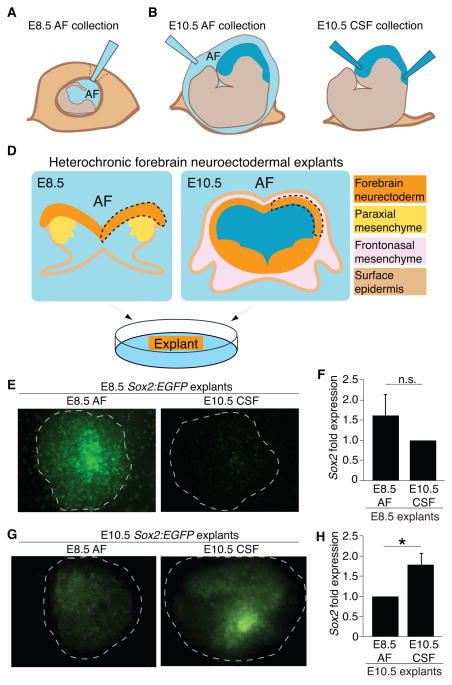
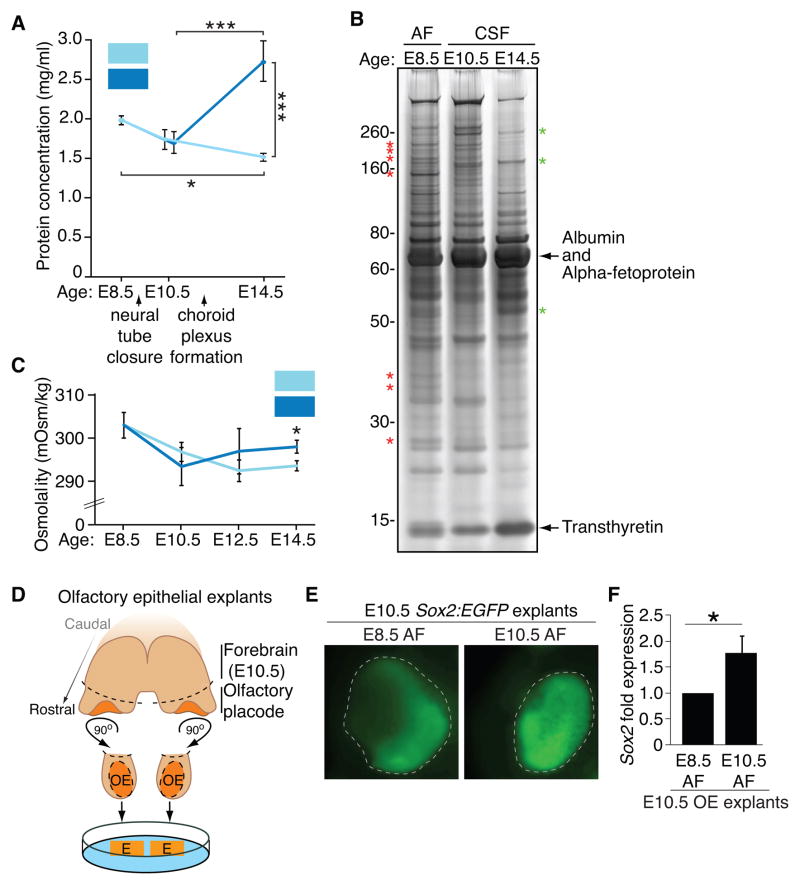
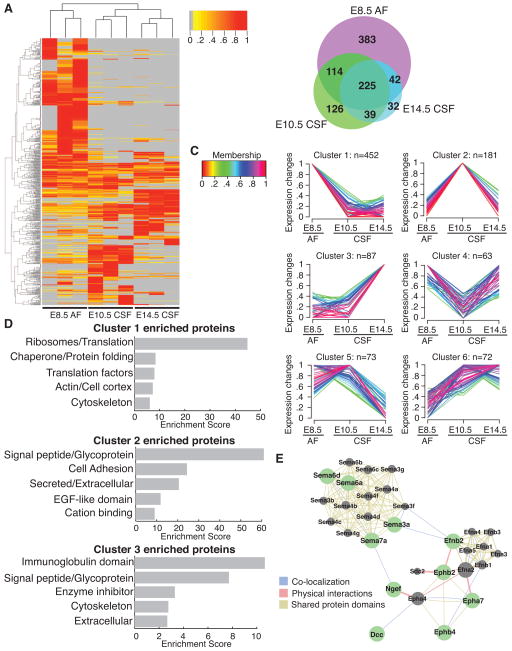
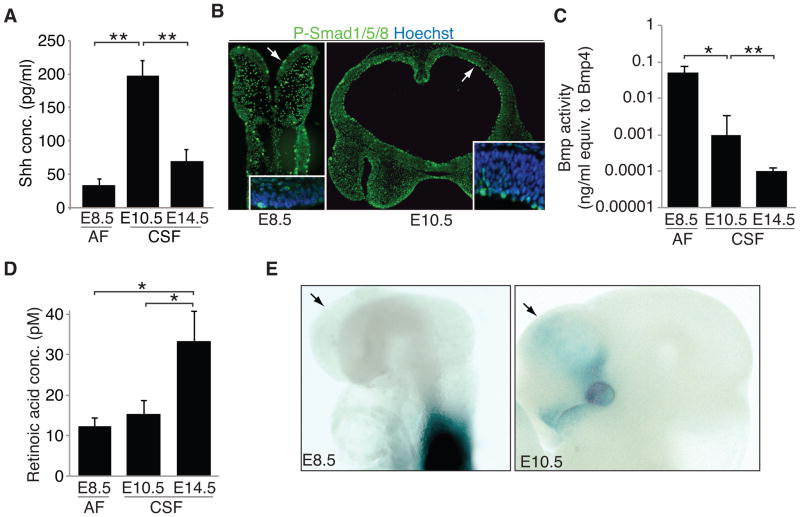
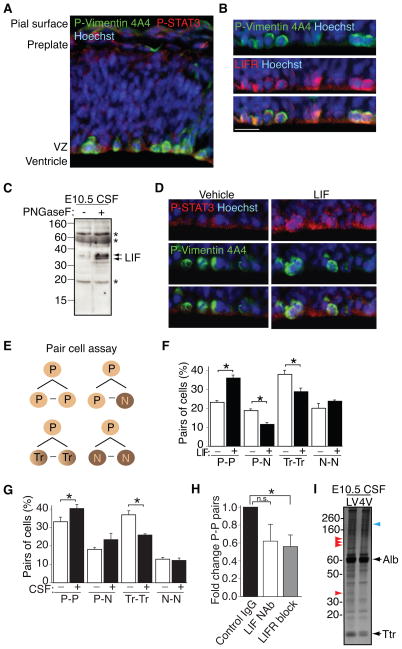
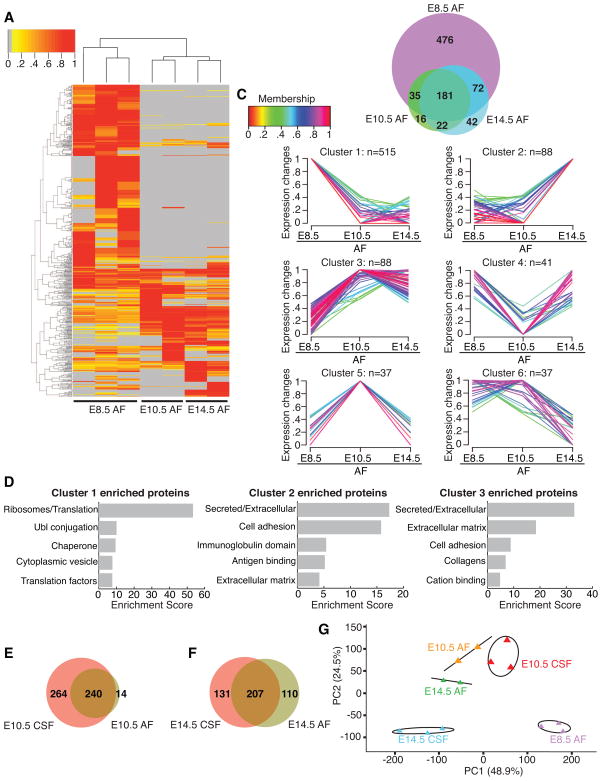
After neural tube closure, amniotic fluid (AF) captured inside the neural tube forms the nascent cerebrospinal fluid (CSF). Neuroepithelial stem cells contact CSF-filled ventricles, proliferate, and differentiate to form the mammalian brain, while neurogenic placodes, which generate cranial sensory neurons, remain in contact with the AF. Using in vivo ultrasound imaging, we quantified the expansion of the embryonic ventricular-CSF space from its inception. We developed tools to obtain pure AF and nascent CSF, before and after neural tube closure, and to define how the AF and CSF proteomes diverge during mouse development. Using embryonic neural explants, we demonstrate that age-matched fluids promote Sox2-positive neurogenic identity in developing forebrain and olfactory epithelia. Nascent CSF also stimulates SOX2-positive self-renewal of forebrain progenitor cells, some of which is attributable to LIFR signaling. Our Resource should facilitate the investigation of fluid-tissue interactions during this highly vulnerable stage of early brain development.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Alonso MI, Gato A, Moro JA, Barbosa E. Disruption of proteoglycans in neural tube fluid by beta-D-xyloside alters brain enlargement in chick embryos. Anat Rec. 1998;252:499–508. - PubMed
-
- Arbeille E, Reynaud F, Sanyas I, Bozon M, Kindbeiter K, Causeret F, Pierani A, Falk J, Moret F, Castellani V. Cerebrospinal fluid-derived Semaphorin3B orients neuroepithelial cell divisions in the apicobasal axis. Nat Commun. 2014;6:6366. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
