

Localized cooling of stems induces latewood formation and cambial dormancy during seasons of active cambium in conifers
- PMID: 26703452
- PMCID: PMC4765539
- DOI: 10.1093/aob/mcv181
Localized cooling of stems induces latewood formation and cambial dormancy during seasons of active cambium in conifers
Abstract
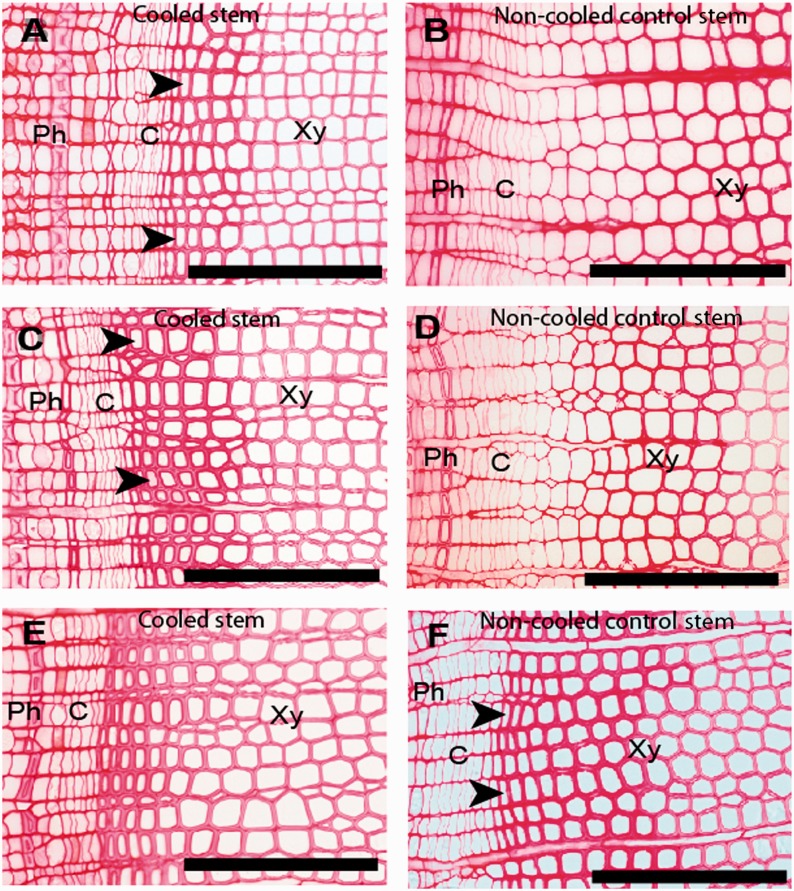
Background and aims: In temperate regions, trees undergo annual cycles of cambial growth, with periods of cambial activity and dormancy. Environmental factors might regulate the cambial growth, as well as the development of cambial derivatives. We investigated the effects of low temperature by localized cooling on cambial activity and latewood formation in two conifers, Chamaecyparis obtusa and Cryptomeria japonica.
Methods: A plastic rubber tube that contained cooled water was wrapped around a 30-cm-wide portion of the main stem of Chamaecyparis obtusa and Cryptomeria japonica trees during seasons of active cambium. Small blocks were collected from both cooled and non-cooled control portions of the stems for sequential observations of cambial activity and for anatomical measurements of cell morphology by light microscopy and image analysis.
Key results: The effect of localized cooling was first observed on differentiating tracheids. Tracheids narrow in diameter and with significantly decreased cambial activity were evident 5 weeks after the start of cooling in these stems. Eight weeks after the start of cooling, tracheids with clearly diminished diameters and thickened cell walls were observed in these stems. Thus, localized low temperature induced narrow diameters and obvious thickening of secondary cell walls of tracheids, which were identified as latewood tracheids. Two months after the cessation of cooling, a false annual ring was observed and cambium became active again and produced new tracheids. In Cryptomeria japonica, cambial activity ceased earlier in locally cooled portions of stems than in non-cooled stems, indicating that the cambium had entered dormancy sooner in the cooled stems.
Conclusions: Artificial cooling of stems induced latewood formation and cessation of cambial activity, indicating that cambium and its derivatives can respond directly to changes in temperature. A decrease in the temperature of the stem is a critical factor in the control of cambial activity and xylem differentiation in trees.
Keywords: Chamaecyparis obtusa; Cryptomeria japonica; cambial dormancy; false ring; latewood formation; localized cooling of stem.
© The Author 2015. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures







References
-
- Barnett JR, Miller H. 1994. The effect of applied heat on graft union formation in dormant Picea sitchensis (Bong.) Carr . Journal of Experimental Botany 45: 135–143.
-
- Begum S, Nakaba S, Bayramzadeh V, Oribe Y, Kubo T, Funada R. 2008. Responses to ambient temperature of cambial reactivation and xylem differentiation in hybrid poplar (Populus sieboldii × P. grandidentata) under natural conditions. Tree Physiology 28: 1813–1819. - PubMed
-
- Begum S, Nakaba S, Oribe Y, Kubo T, Funada R. 2010a. Cambial sensitivity to rising temperatures from late winter to early spring in the evergreen conifer Cryptomeria japonica . Trees 24: 43–52.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
