

R-Ras Regulates Murine T Cell Migration and Intercellular Adhesion Molecule-1 Binding
- PMID: 26710069
- PMCID: PMC4692399
- DOI: 10.1371/journal.pone.0145218
R-Ras Regulates Murine T Cell Migration and Intercellular Adhesion Molecule-1 Binding
Abstract
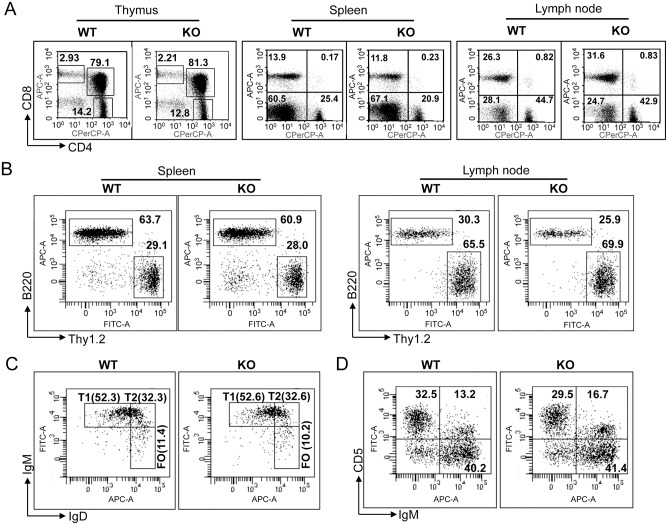
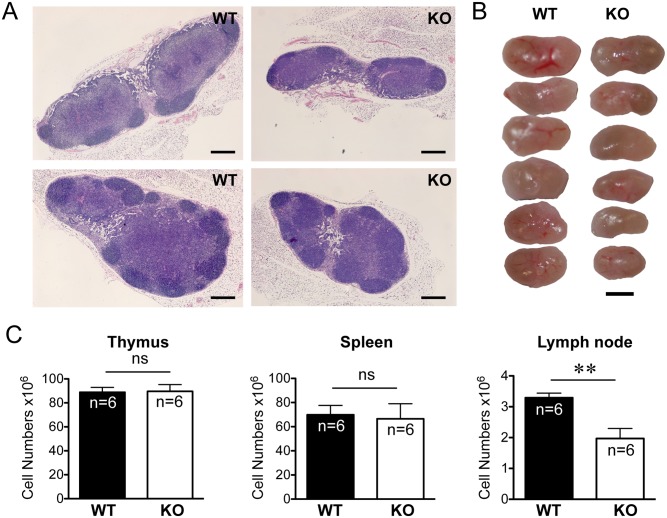
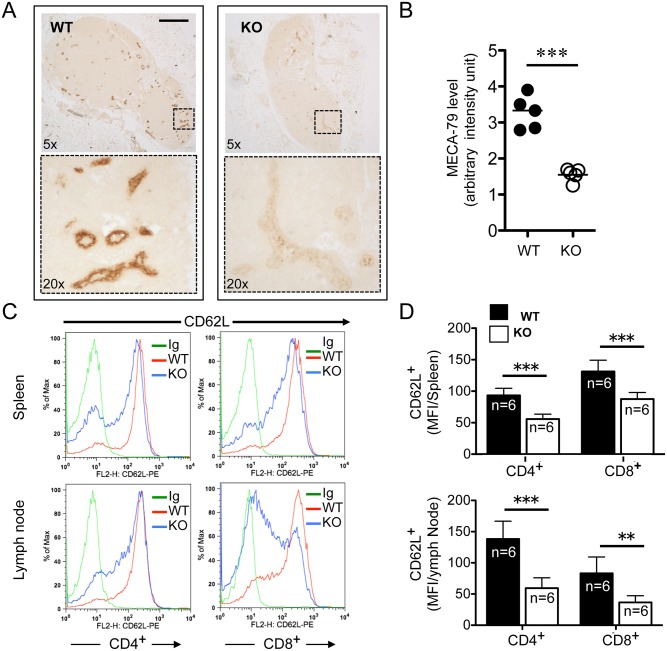
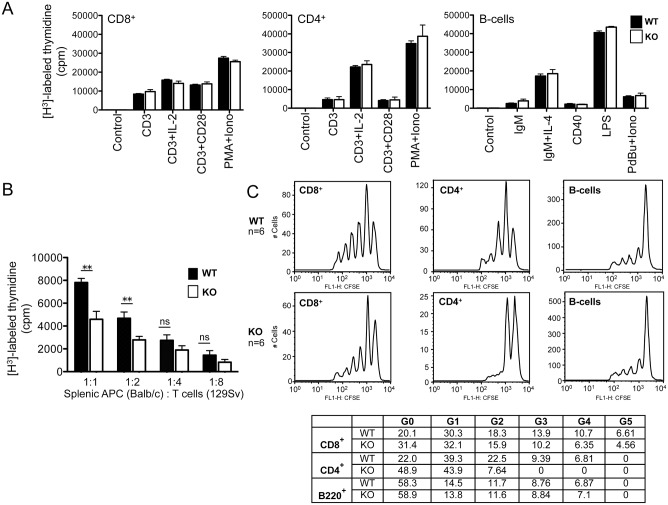
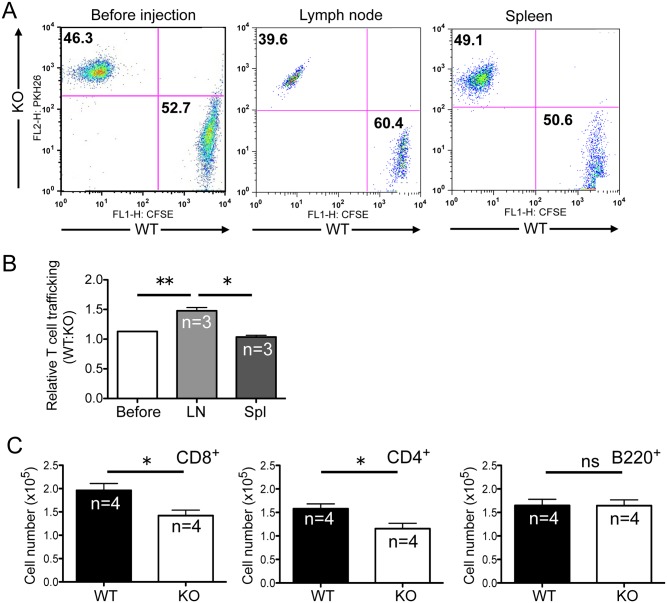
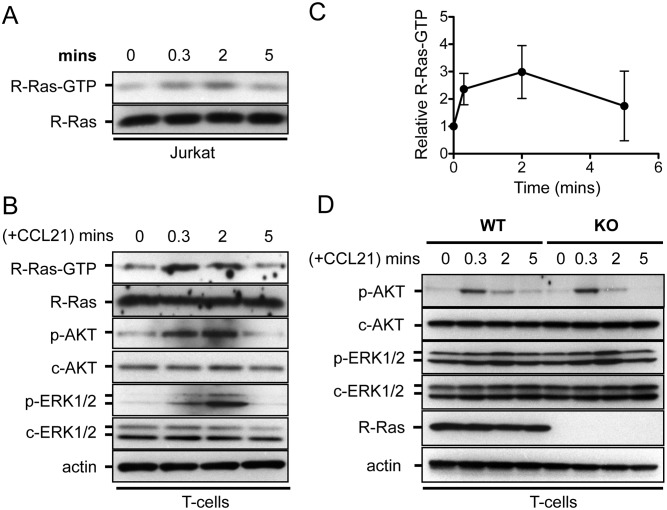
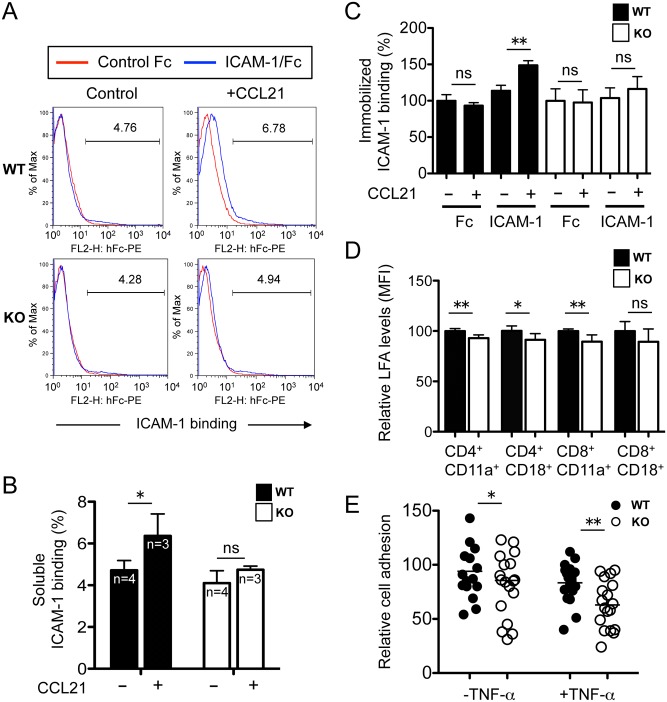
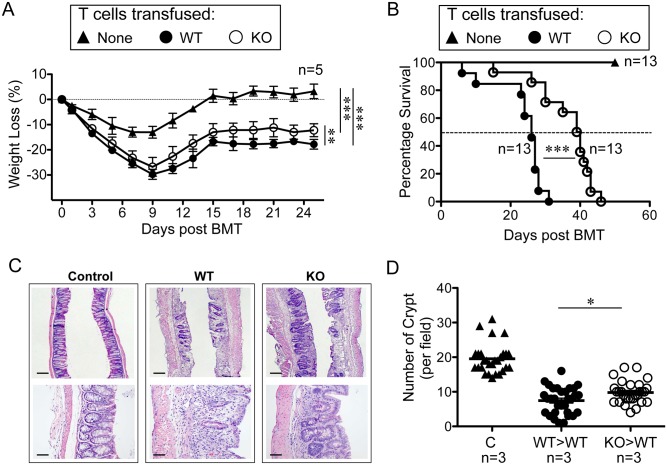
The trafficking of T-lymphocytes to peripheral draining lymph nodes is crucial for mounting an adaptive immune response. The role of chemokines in the activation of integrins via Ras-related small GTPases has been well established. R-Ras is a member of the Ras-subfamily of small guanosine-5'-triphosphate-binding proteins and its role in T cell trafficking has been investigated in R-Ras null mice (Rras-/-). An examination of the lymphoid organs of Rras-/- mice revealed a 40% reduction in the cellularity of the peripheral lymph nodes. Morphologically, the high endothelial venules of Rras-/- mice were more disorganized and less mature than those of wild-type mice. Furthermore, CD4+ and CD8+ T cells from Rras-/- mice had approximately 42% lower surface expression of L-selectin/CD62L. These aberrant peripheral lymph node phenotypes were associated with proliferative and trafficking defects in Rras-/- T cells. Furthermore, R-Ras could be activated by the chemokine, CCL21. Indeed, Rras-/- T cells had approximately 14.5% attenuation in binding to intercellular adhesion molecule 1 upon CCL21 stimulation. Finally, in a graft-versus host disease model, recipient mice that were transfused with Rras-/- T cells showed a significant reduction in disease severity when compared with mice transplanted with wild-type T cells. These findings implicate a role for R-Ras in T cell trafficking in the high endothelial venules during an effective immune response.
Conflict of interest statement
Figures
References
-
- Saez R, Chan AM, Miki T, Aaronson SA. Oncogenic activation of human R-ras by point mutations analogous to those of prototype H-ras oncogenes. Oncogene. 1994;9(10):2977–82. Epub 1994/10/01. . - PubMed
-
- Cox AD, Brtva TR, Lowe DG, Der CJ. R-Ras induces malignant, but not morphologic, transformation of NIH3T3 cells. Oncogene. 1994;9(11):3281–8. Epub 1994/11/01. . - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U54 HL090503/HL/NHLBI NIH HHS/United States
- HL090503/HL/NHLBI NIH HHS/United States
- P01 HL044612/HL/NHLBI NIH HHS/United States
- HL102836/HL/NHLBI NIH HHS/United States
- R01 HL102836/HL/NHLBI NIH HHS/United States
- CA78509/CA/NCI NIH HHS/United States
- R01 AI079087/AI/NIAID NIH HHS/United States
- R01 MH059771/MH/NIMH NIH HHS/United States
- HL44612/HL/NHLBI NIH HHS/United States
- AI079087/AI/NIAID NIH HHS/United States
- MH59771/MH/NIMH NIH HHS/United States
- T32 CA078207/CA/NCI NIH HHS/United States
- R01 NS070711/NS/NINDS NIH HHS/United States
- NS00711/NS/NINDS NIH HHS/United States
- T32CA078207/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
