

Phosphorylation of Ku dictates DNA double-strand break (DSB) repair pathway choice in S phase
- PMID: 26712563
- PMCID: PMC4770226
- DOI: 10.1093/nar/gkv1499
Phosphorylation of Ku dictates DNA double-strand break (DSB) repair pathway choice in S phase
Abstract
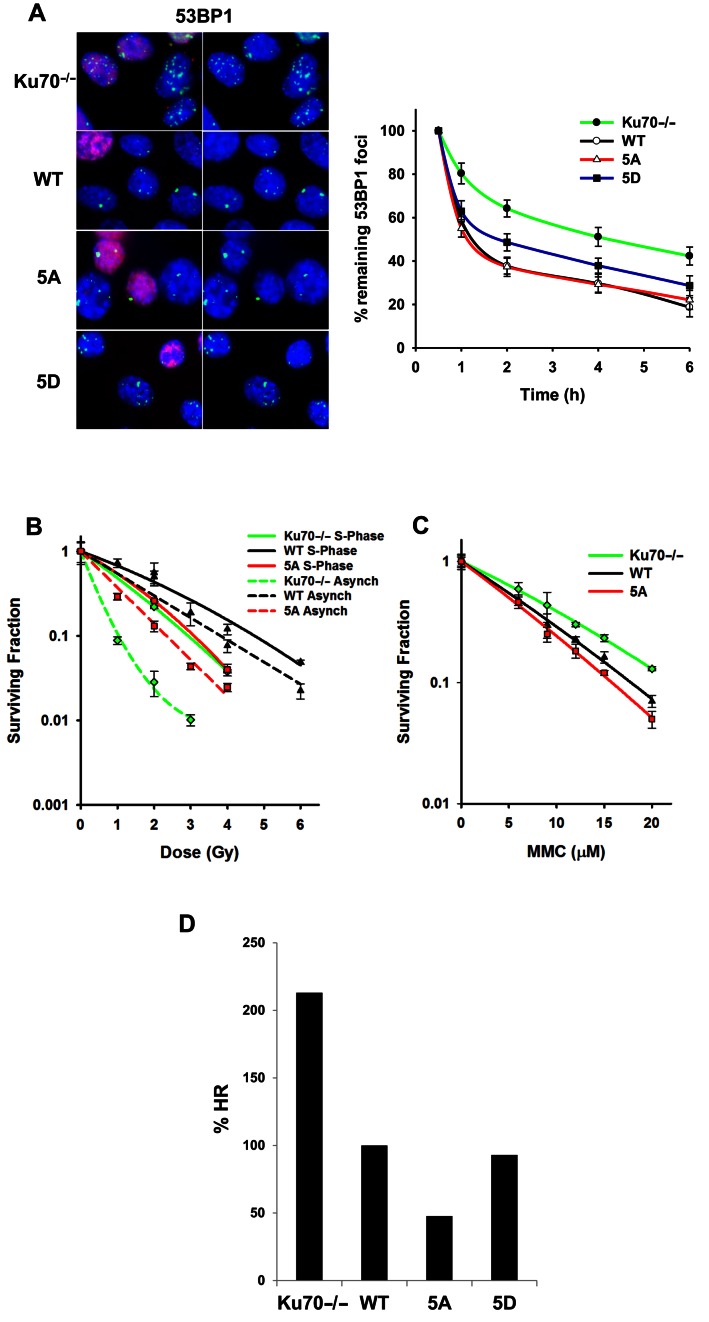
Multiple DNA double-strand break (DSB) repair pathways are active in S phase of the cell cycle; however, DSBs are primarily repaired by homologous recombination (HR) in this cell cycle phase. As the non-homologous end-joining (NHEJ) factor, Ku70/80 (Ku), is quickly recruited to DSBs in S phase, we hypothesized that an orchestrated mechanism modulates pathway choice between HR and NHEJ via displacement of the Ku heterodimer from DSBs to allow HR. Here, we provide evidence that phosphorylation at a cluster of sites in the junction of the pillar and bridge regions of Ku70 mediates the dissociation of Ku from DSBs. Mimicking phosphorylation at these sites reduces Ku's affinity for DSB ends, suggesting that phosphorylation of Ku70 induces a conformational change responsible for the dissociation of the Ku heterodimer from DNA ends. Ablating phosphorylation of Ku70 leads to the sustained retention of Ku at DSBs, resulting in a significant decrease in DNA end resection and HR, specifically in S phase. This decrease in HR is specific as these phosphorylation sites are not required for NHEJ. Our results demonstrate that the phosphorylation-mediated dissociation of Ku70/80 from DSBs frees DNA ends, allowing the initiation of HR in S phase and providing a mechanism of DSB repair pathway choice in mammalian cells.
© The Author(s) 2015. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
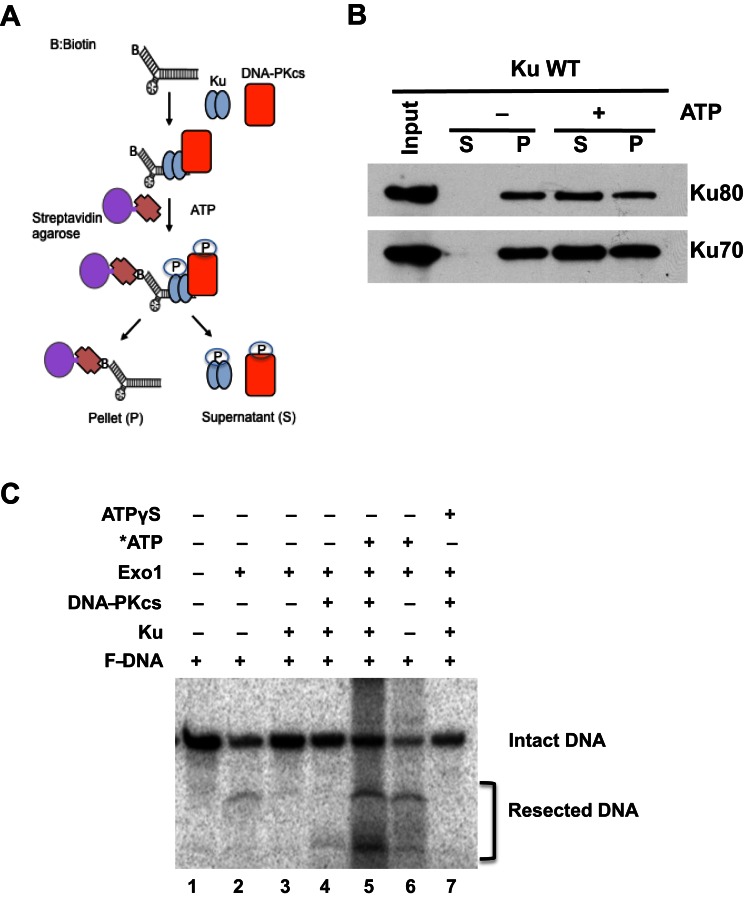
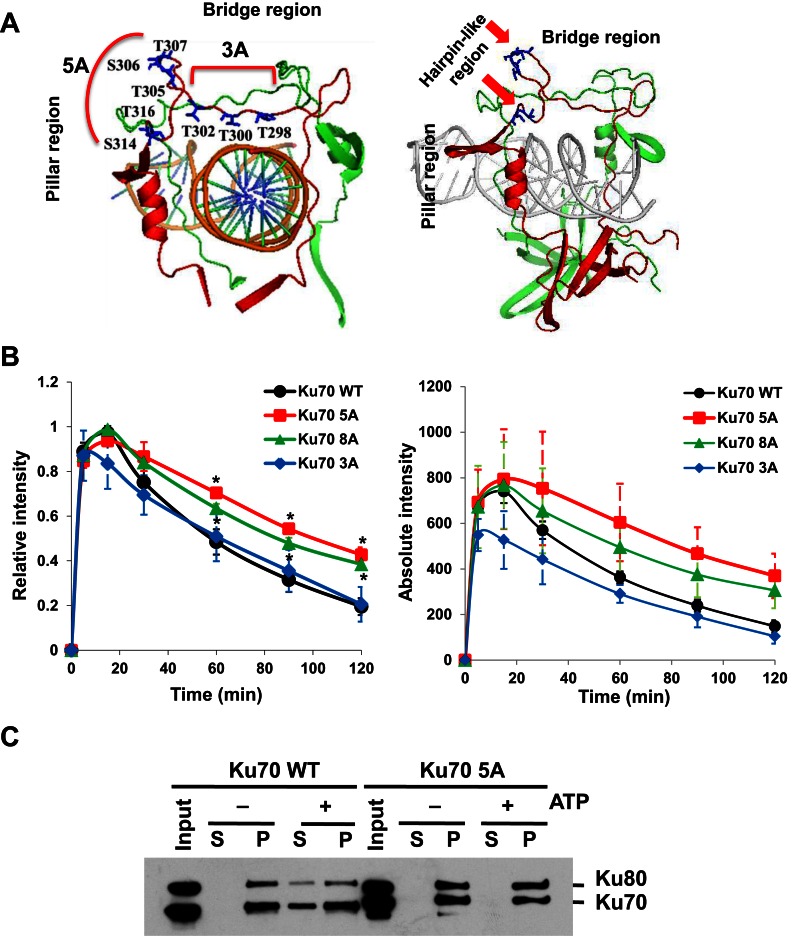
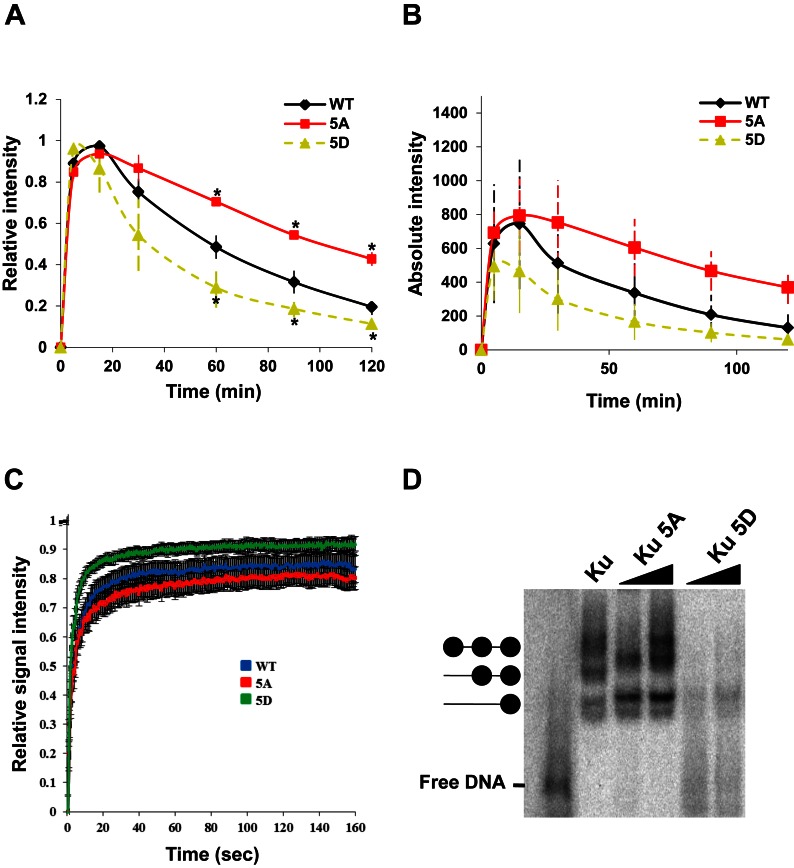
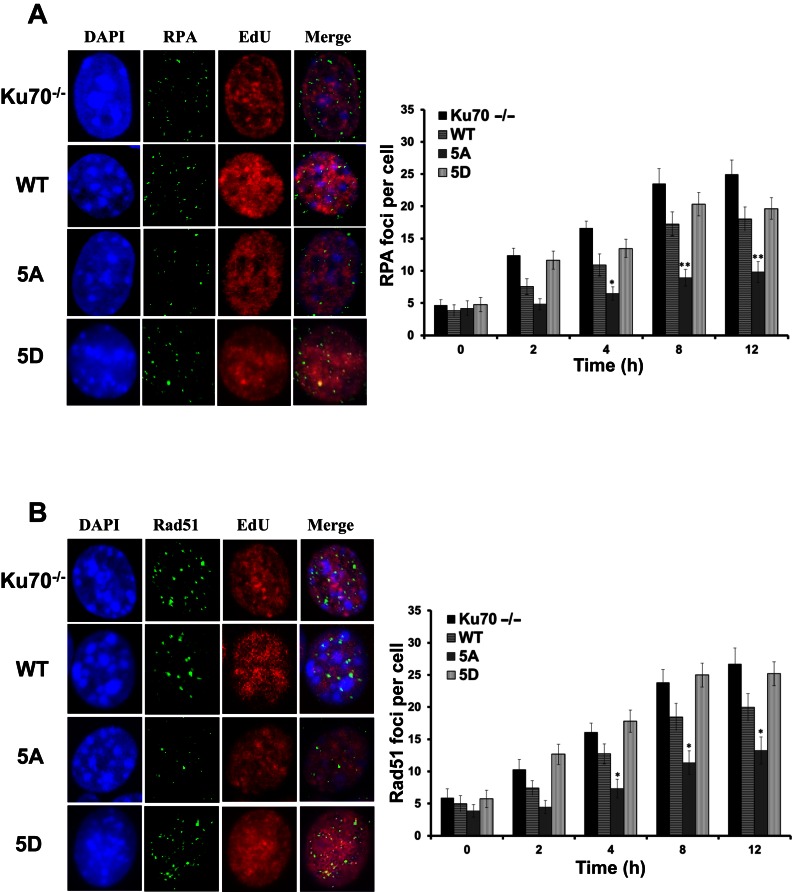
References
-
- Hoeijmakers J.H. Genome maintenance mechanisms for preventing cancer. Nature. 2001;411:366–374. - PubMed
-
- Goodarzi A.A., Jeggo P.A. The repair and signaling responses to DNA double-strand breaks. Adv. Genet. 2013;82:1–45. - PubMed
-
- Thompson L.H. Recognition, signaling, and repair of DNA double-strand breaks produced by ionizing radiation in mammalian cells: the molecular choreography. Mutat. Res. 2012;751:158–246. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
