

The hitchhiker's guide to the voltage-gated sodium channel galaxy
- PMID: 26712848
- PMCID: PMC4692491
- DOI: 10.1085/jgp.201511492
The hitchhiker's guide to the voltage-gated sodium channel galaxy
Abstract
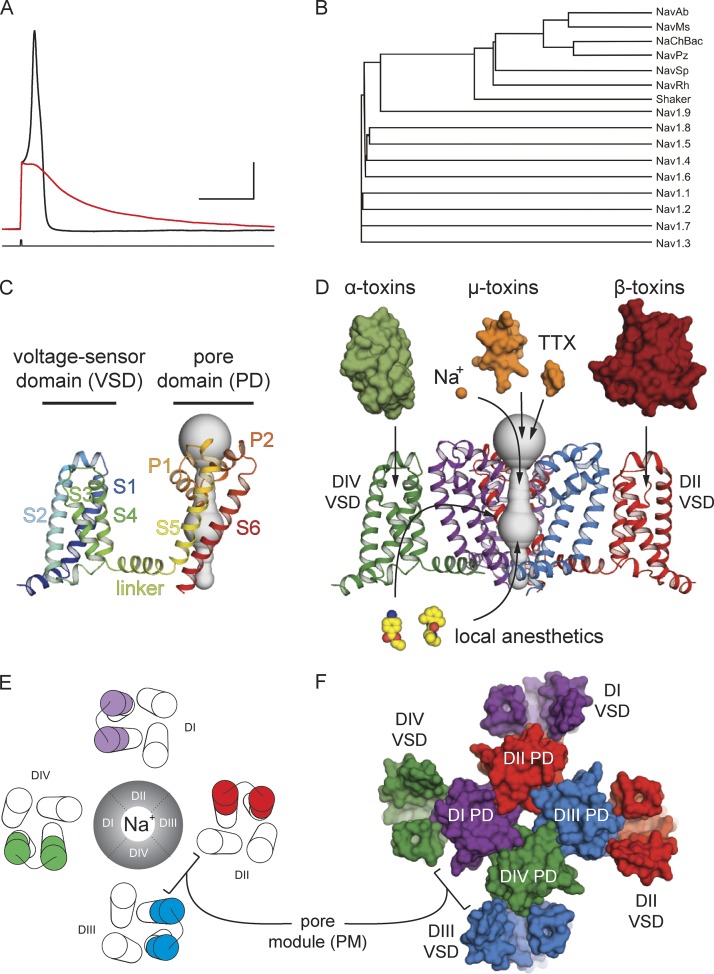
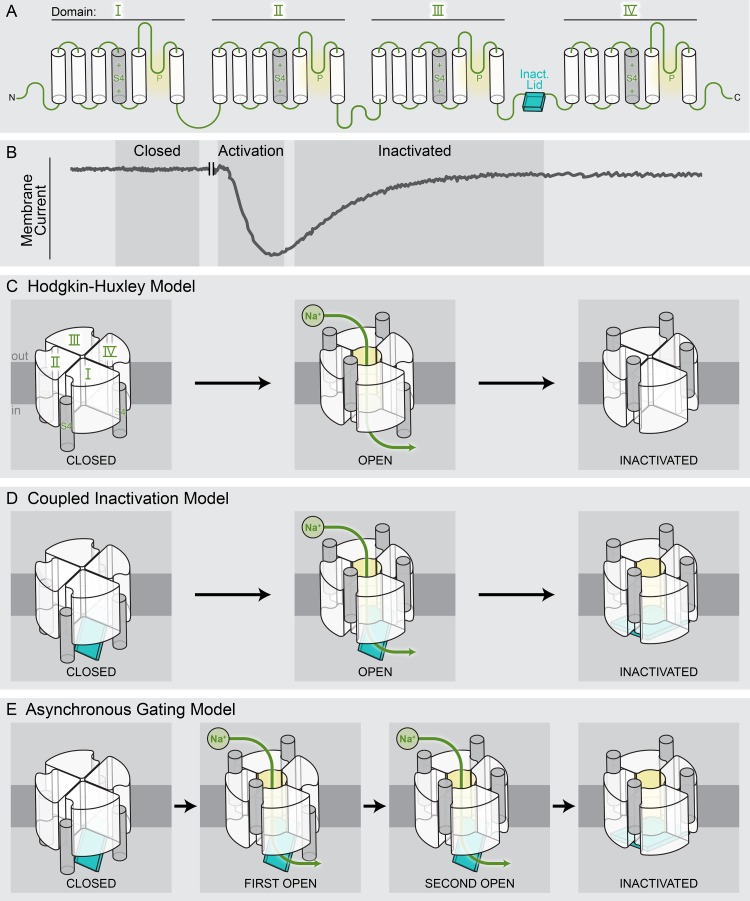
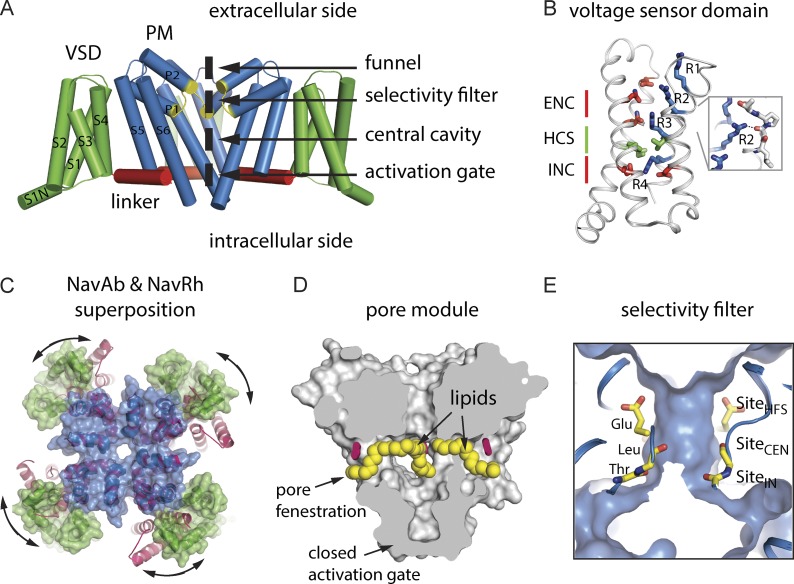
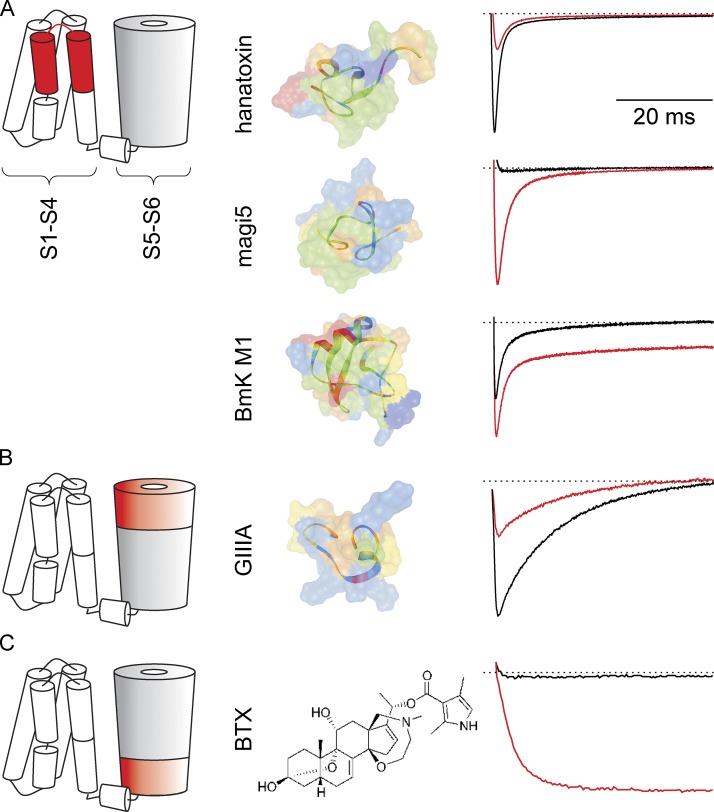
Eukaryotic voltage-gated sodium (Nav) channels contribute to the rising phase of action potentials and served as an early muse for biophysicists laying the foundation for our current understanding of electrical signaling. Given their central role in electrical excitability, it is not surprising that (a) inherited mutations in genes encoding for Nav channels and their accessory subunits have been linked to excitability disorders in brain, muscle, and heart; and (b) Nav channels are targeted by various drugs and naturally occurring toxins. Although the overall architecture and behavior of these channels are likely to be similar to the more well-studied voltage-gated potassium channels, eukaryotic Nav channels lack structural and functional symmetry, a notable difference that has implications for gating and selectivity. Activation of voltage-sensing modules of the first three domains in Nav channels is sufficient to open the channel pore, whereas movement of the domain IV voltage sensor is correlated with inactivation. Also, structure-function studies of eukaryotic Nav channels show that a set of amino acids in the selectivity filter, referred to as DEKA locus, is essential for Na(+) selectivity. Structures of prokaryotic Nav channels have also shed new light on mechanisms of drug block. These structures exhibit lateral fenestrations that are large enough to allow drugs or lipophilic molecules to gain access into the inner vestibule, suggesting that this might be the passage for drug entry into a closed channel. In this Review, we will synthesize our current understanding of Nav channel gating mechanisms, ion selectivity and permeation, and modulation by therapeutics and toxins in light of the new structures of the prokaryotic Nav channels that, for the time being, serve as structural models of their eukaryotic counterparts.
© 2016 Ahern et al.
Figures
References
-
- Agnew W.S., Levinson S.R., Brabson J.S., and Raftery M.A.. 1978. Purification of the tetrodotoxin-binding component associated with the voltage-sensitive sodium channel from Electrophorus electricus electroplax membranes. Proc. Natl. Acad. Sci. USA. 75:2606–2610. 10.1073/pnas.75.6.2606 - DOI - PMC - PubMed
-
- Ahuja S., Mukund S., Deng L., Khakh K., Chang E., Ho H., Shriver S., Young C., Lin S., Johnson J.P. Jr., et al. 2015. Structural basis of Nav1.7 inhibition by an isoform-selective small molecule antagonist. Science. 350:1491. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
