

Expression of the FGFR2 mesenchymal splicing variant in epithelial cells drives epithelial-mesenchymal transition
- PMID: 26713601
- PMCID: PMC4868697
- DOI: 10.18632/oncotarget.6706
Expression of the FGFR2 mesenchymal splicing variant in epithelial cells drives epithelial-mesenchymal transition
Abstract
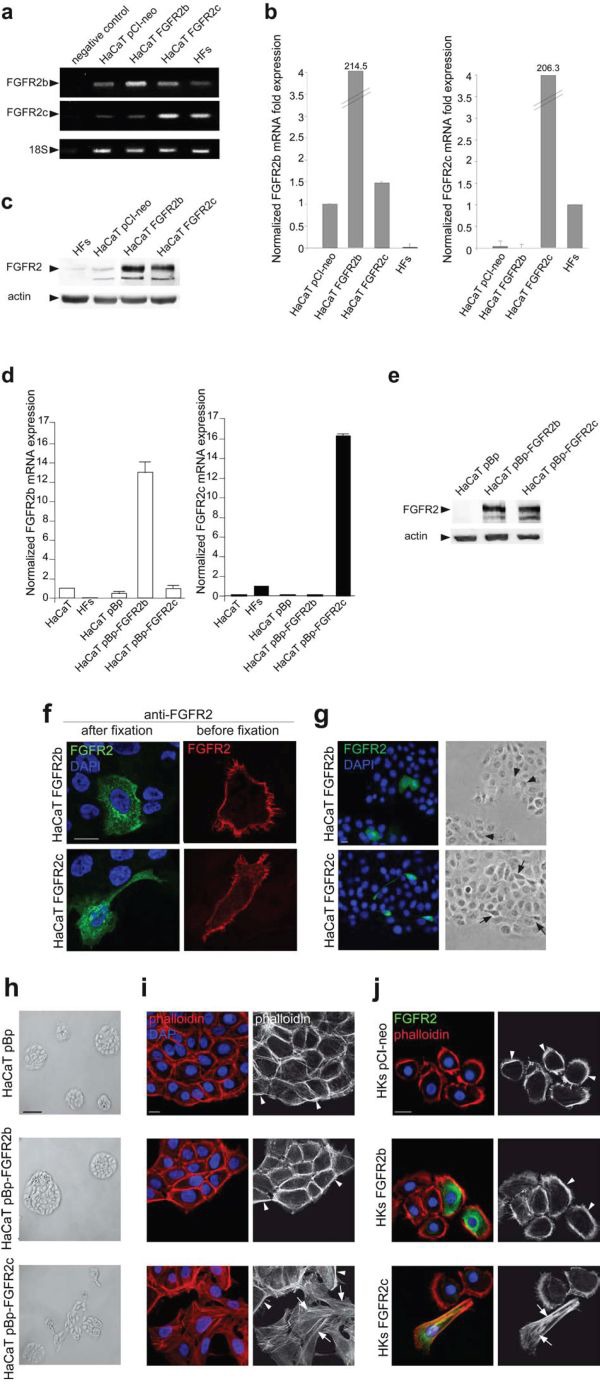
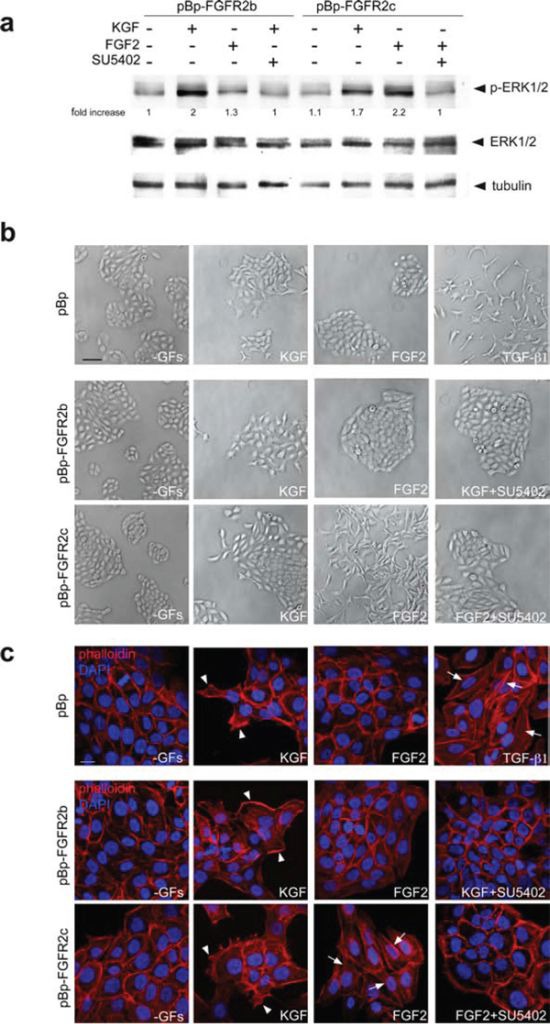
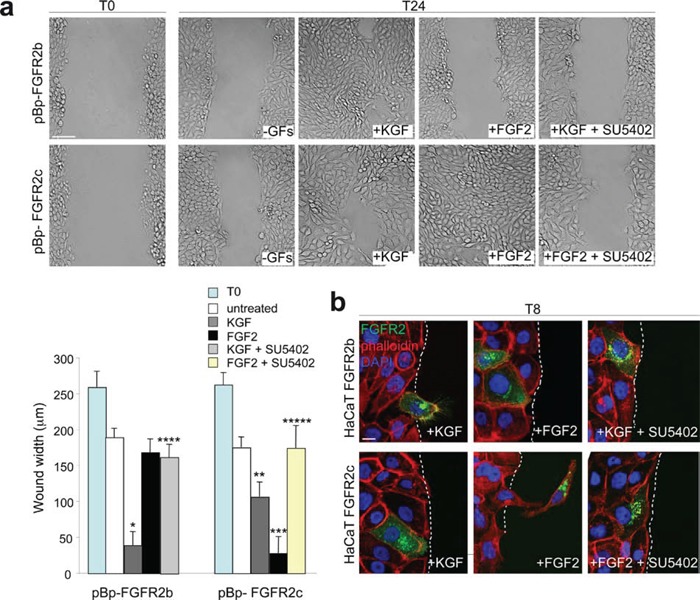
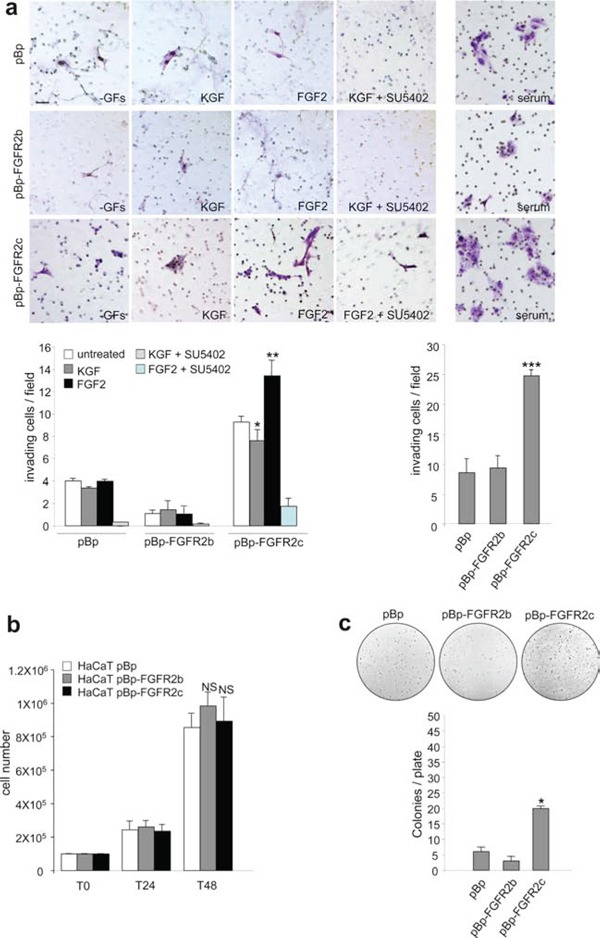
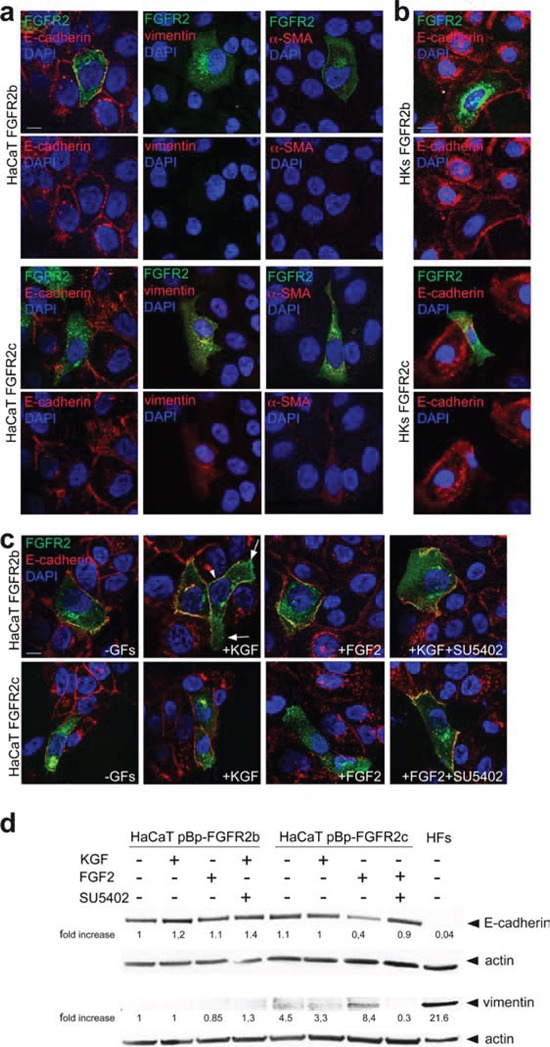
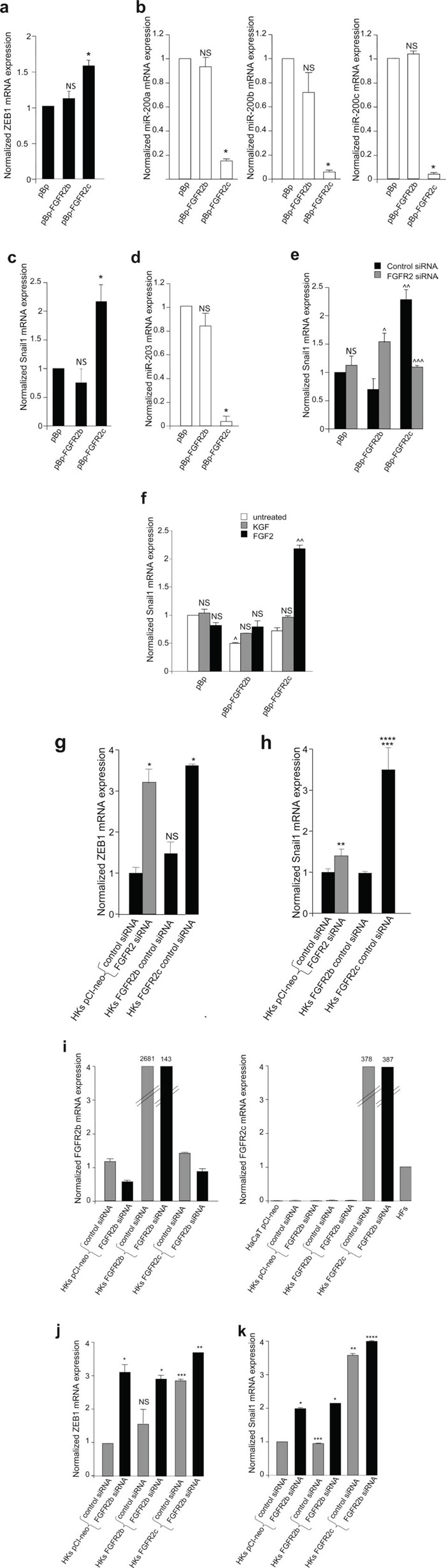
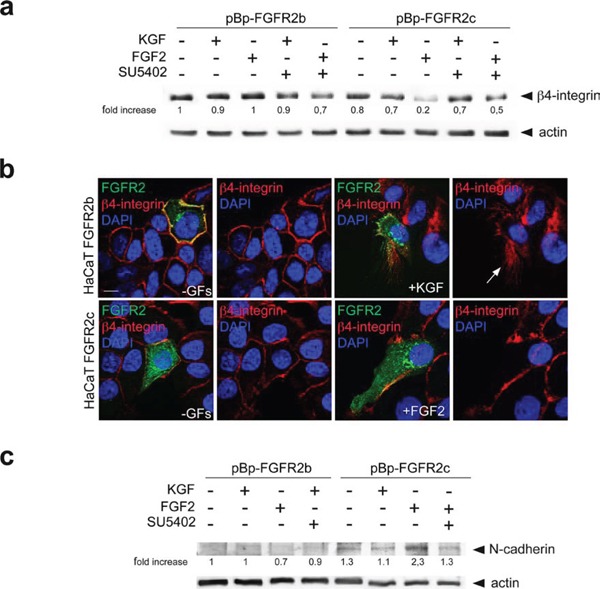
The FGFRs are receptor tyrosine kinases expressed by tissue-specific alternative splicing in epithelial IIIb or mesenchymal IIIc isoforms. Deregulation of FGF/FGFR signaling unbalances the epithelial-stromal homeostasis and may lead to cancer development. In the epithelial-context, while FGFR2b/KGFR acts as tumor suppressor, FGFR2c appears to play an oncogenic role. Based on our recent observation that the switching of FGFR2b versus FGFR2c induces EMT, here we investigated the biological outcome of the ectopic expression of FGFR2c in normal human keratinocytes. Morphological analysis showed that, differently from FGFR2b overexpression, the forced expression and activation of FGFR2c drive the epithelial cells to acquire a mesenchymal-like shape and actin reorganization. Moreover, the appearance of invasiveness and anchorage-independent growth ability in FGFR2c transfected keratinocytes was consistent with the potential tumorigenic role proposed for this receptor variant. Biochemical and molecular approaches revealed that the observed phenotypic changes were accompanied by modulation of EMT biomarkers and indicated the involvement of EMT transcription factors and miRs. Finally, the analysis of the expression pattern of discriminating markers strongly suggested that activation of FGFR2c triggers a process corresponding to the initiation of the pathological type III EMT, but not to the more physiological type II EMT occurring during FGFR2b-mediated wound healing.
Keywords: FGFR2; epithelial-mesenchymal transition; human keratinocytes.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
Expression of the FGFR2c mesenchymal splicing variant in human keratinocytes inhibits differentiation and promotes invasion.Mol Carcinog. 2018 Feb;57(2):272-283. doi: 10.1002/mc.22754. Epub 2017 Nov 14. Mol Carcinog. 2018. PMID: 29068468 Free PMC article.
-
The aberrant expression in epithelial cells of the mesenchymal isoform of FGFR2 controls the negative crosstalk between EMT and autophagy.J Cell Mol Med. 2021 Apr;25(8):4166-4172. doi: 10.1111/jcmm.16309. Epub 2021 Feb 20. J Cell Mol Med. 2021. PMID: 33609078 Free PMC article.
-
HPV16 E5 expression induces switching from FGFR2b to FGFR2c and epithelial-mesenchymal transition.Int J Cancer. 2015 Jul 1;137(1):61-72. doi: 10.1002/ijc.29373. Epub 2014 Dec 13. Int J Cancer. 2015. PMID: 25450802
-
FGFR2-related pathogenesis and FGFR2-targeted therapeutics (Review).Int J Mol Med. 2009 Mar;23(3):307-11. doi: 10.3892/ijmm_00000132. Int J Mol Med. 2009. PMID: 19212647 Review.
-
Role of fibroblast growth factor receptor-2 splicing in normal and cancer cells.Front Biosci (Landmark Ed). 2018 Jan 1;23(4):626-639. doi: 10.2741/4609. Front Biosci (Landmark Ed). 2018. PMID: 28930565 Review.
Cited by
-
Expression Profile of Fibroblast Growth Factor Receptors, Keratinocyte Differentiation Markers, and Epithelial Mesenchymal Transition-Related Genes in Actinic Keratosis: A Possible Predictive Factor for Malignant Progression?Biology (Basel). 2021 Apr 15;10(4):331. doi: 10.3390/biology10040331. Biology (Basel). 2021. PMID: 33920760 Free PMC article.
-
Epithelial splicing regulatory protein 1 and 2 paralogues correlate with splice signatures and favorable outcome in human colorectal cancer.Oncotarget. 2016 Nov 8;7(45):73800-73816. doi: 10.18632/oncotarget.12070. Oncotarget. 2016. PMID: 27650542 Free PMC article.
-
Hyaluronic Acid (HA), Platelet-Rich Plasm and Extracorporeal Shock Wave Therapy (ESWT) promote human chondrocyte regeneration in vitro and ESWT-mediated increase of CD44 expression enhances their susceptibility to HA treatment.PLoS One. 2019 Jun 28;14(6):e0218740. doi: 10.1371/journal.pone.0218740. eCollection 2019. PLoS One. 2019. PMID: 31251756 Free PMC article.
-
FGFR2c Upregulation Contributes to Cancer-Associated Fibroblast Program Activation and to Enhanced Autophagy in Actinic Keratosis-Derived Dermal Fibroblasts: A Possible Role in Precancerous Cell/Stromal Cell Crosstalk.Biology (Basel). 2023 Mar 16;12(3):463. doi: 10.3390/biology12030463. Biology (Basel). 2023. PMID: 36979155 Free PMC article.
-
Role of PKCε in the epithelial-mesenchymal transition induced by FGFR2 isoform switch.Cell Commun Signal. 2020 May 19;18(1):76. doi: 10.1186/s12964-020-00582-1. Cell Commun Signal. 2020. PMID: 32429937 Free PMC article.
References
-
- Turner N, Grose R. Fibroblast growth factor signalling: from development to cancer. Nat Rev Cancer. 2010;10:116–129. - PubMed
-
- Haugsten EM, Wiedlocha A, Olsnes S, Wesche J. Roles of fibroblast growth factor receptors in cancerogenesis. Mol Cancer Res. 2010;8:1439–1452. - PubMed
-
- Brooks AN, Kilgour E, Smith PD. Molecular pathways: fibroblast growth factor signaling: a new therapeutic opportunity in cancer. Clin Cancer Res. 2012;18:1855–1862. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
