

Where's the Noise? Key Features of Spontaneous Activity and Neural Variability Arise through Learning in a Deterministic Network
- PMID: 26714277
- PMCID: PMC4694925
- DOI: 10.1371/journal.pcbi.1004640
Where's the Noise? Key Features of Spontaneous Activity and Neural Variability Arise through Learning in a Deterministic Network
Abstract
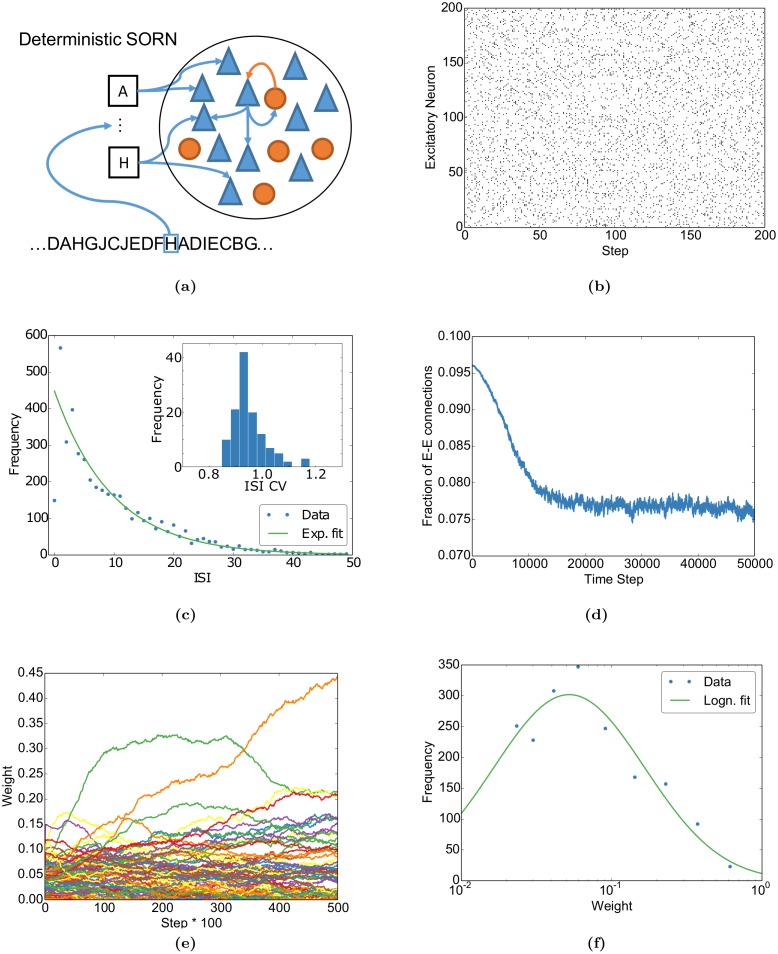
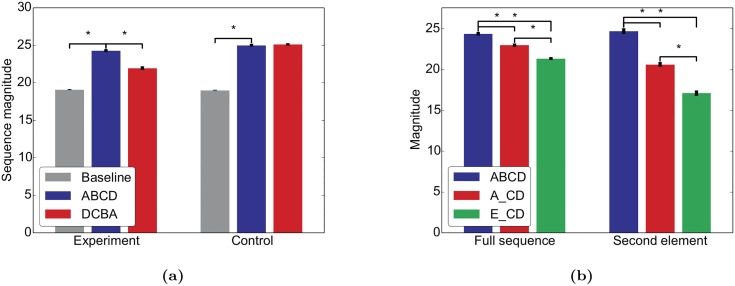
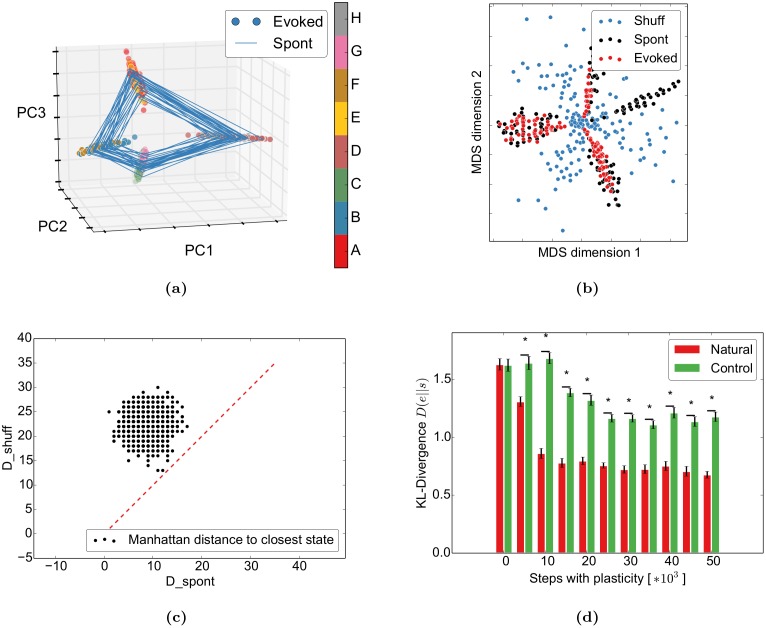
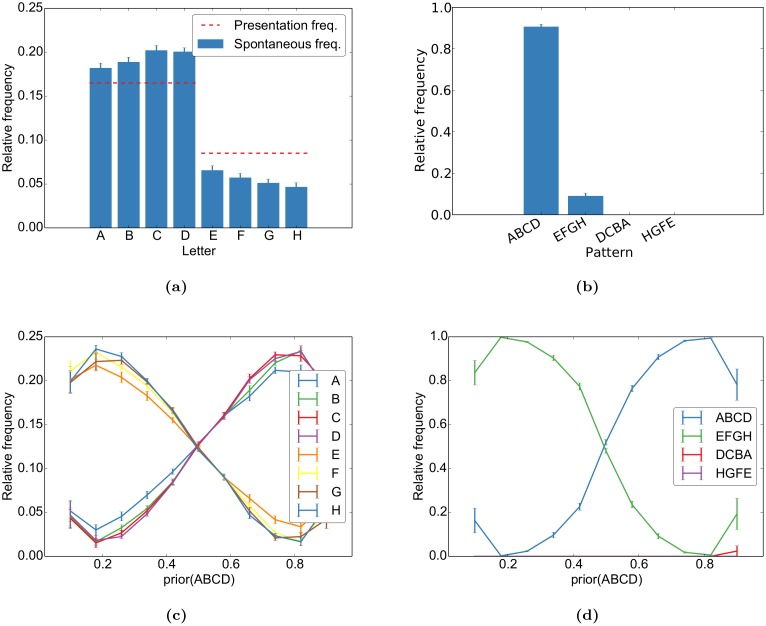
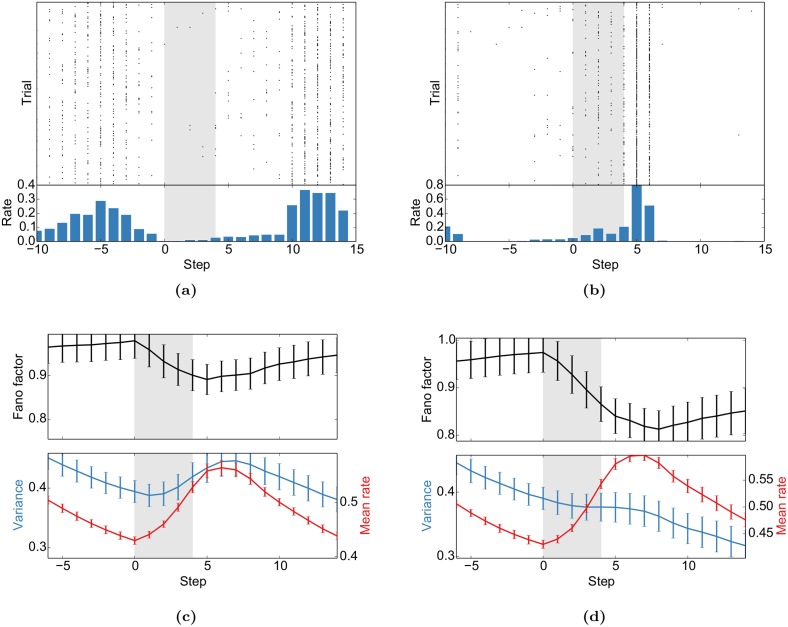
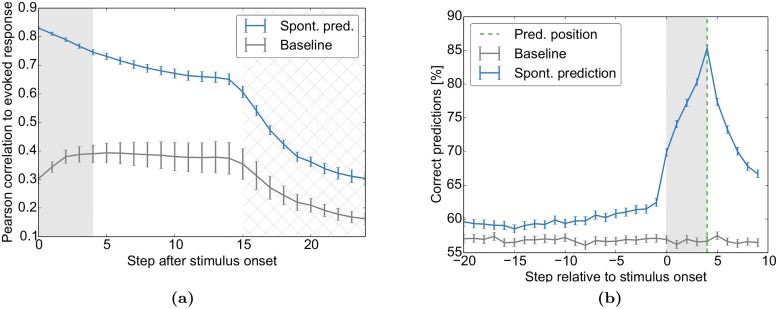
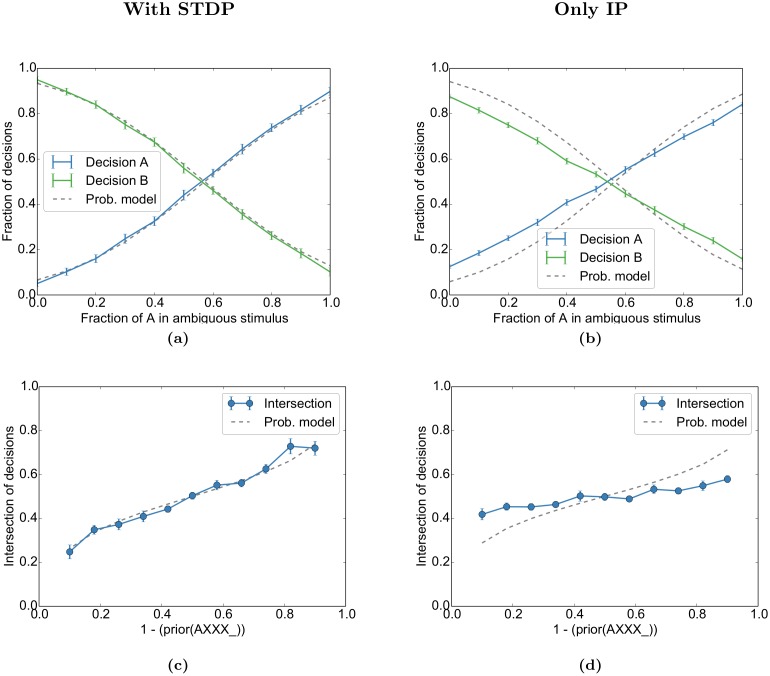
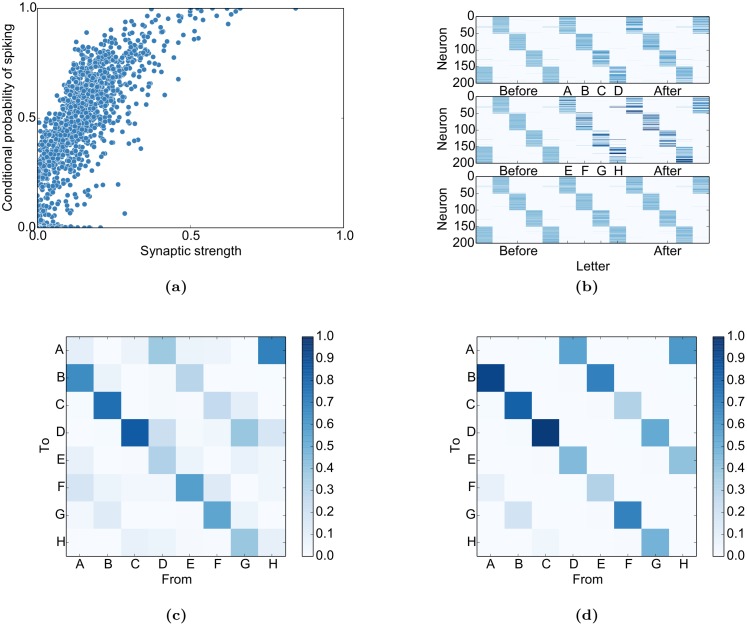
Even in the absence of sensory stimulation the brain is spontaneously active. This background "noise" seems to be the dominant cause of the notoriously high trial-to-trial variability of neural recordings. Recent experimental observations have extended our knowledge of trial-to-trial variability and spontaneous activity in several directions: 1. Trial-to-trial variability systematically decreases following the onset of a sensory stimulus or the start of a motor act. 2. Spontaneous activity states in sensory cortex outline the region of evoked sensory responses. 3. Across development, spontaneous activity aligns itself with typical evoked activity patterns. 4. The spontaneous brain activity prior to the presentation of an ambiguous stimulus predicts how the stimulus will be interpreted. At present it is unclear how these observations relate to each other and how they arise in cortical circuits. Here we demonstrate that all of these phenomena can be accounted for by a deterministic self-organizing recurrent neural network model (SORN), which learns a predictive model of its sensory environment. The SORN comprises recurrently coupled populations of excitatory and inhibitory threshold units and learns via a combination of spike-timing dependent plasticity (STDP) and homeostatic plasticity mechanisms. Similar to balanced network architectures, units in the network show irregular activity and variable responses to inputs. Additionally, however, the SORN exhibits sequence learning abilities matching recent findings from visual cortex and the network's spontaneous activity reproduces the experimental findings mentioned above. Intriguingly, the network's behaviour is reminiscent of sampling-based probabilistic inference, suggesting that correlates of sampling-based inference can develop from the interaction of STDP and homeostasis in deterministic networks. We conclude that key observations on spontaneous brain activity and the variability of neural responses can be accounted for by a simple deterministic recurrent neural network which learns a predictive model of its sensory environment via a combination of generic neural plasticity mechanisms.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
A model of human motor sequence learning explains facilitation and interference effects based on spike-timing dependent plasticity.PLoS Comput Biol. 2017 Aug 2;13(8):e1005632. doi: 10.1371/journal.pcbi.1005632. eCollection 2017 Aug. PLoS Comput Biol. 2017. PMID: 28767646 Free PMC article.
-
A learning theory for reward-modulated spike-timing-dependent plasticity with application to biofeedback.PLoS Comput Biol. 2008 Oct;4(10):e1000180. doi: 10.1371/journal.pcbi.1000180. Epub 2008 Oct 10. PLoS Comput Biol. 2008. PMID: 18846203 Free PMC article.
-
Bridging structure and function: A model of sequence learning and prediction in primary visual cortex.PLoS Comput Biol. 2018 Jun 5;14(6):e1006187. doi: 10.1371/journal.pcbi.1006187. eCollection 2018 Jun. PLoS Comput Biol. 2018. PMID: 29870532 Free PMC article.
-
A theory of cortical responses.Philos Trans R Soc Lond B Biol Sci. 2005 Apr 29;360(1456):815-36. doi: 10.1098/rstb.2005.1622. Philos Trans R Soc Lond B Biol Sci. 2005. PMID: 15937014 Free PMC article. Review.
-
The Brain as an Efficient and Robust Adaptive Learner.Neuron. 2017 Jun 7;94(5):969-977. doi: 10.1016/j.neuron.2017.05.016. Neuron. 2017. PMID: 28595053 Review.
Cited by
-
Multiple states in ongoing neural activity in the rat visual cortex.PLoS One. 2021 Aug 26;16(8):e0256791. doi: 10.1371/journal.pone.0256791. eCollection 2021. PLoS One. 2021. PMID: 34437630 Free PMC article.
-
Neural Code-Neural Self-information Theory on How Cell-Assembly Code Rises from Spike Time and Neuronal Variability.Front Cell Neurosci. 2017 Aug 30;11:236. doi: 10.3389/fncel.2017.00236. eCollection 2017. Front Cell Neurosci. 2017. PMID: 28912685 Free PMC article.
-
Overexpression of cypin alters dendrite morphology, single neuron activity, and network properties via distinct mechanisms.J Neural Eng. 2018 Feb;15(1):016020. doi: 10.1088/1741-2552/aa976a. J Neural Eng. 2018. PMID: 29091046 Free PMC article.
-
Robust encoding of natural stimuli by neuronal response sequences in monkey visual cortex.Nat Commun. 2023 May 25;14(1):3021. doi: 10.1038/s41467-023-38587-2. Nat Commun. 2023. PMID: 37231014 Free PMC article.
-
Cortical Variability and Challenges for Modeling Approaches.Front Syst Neurosci. 2017 Apr 4;11:15. doi: 10.3389/fnsys.2017.00015. eCollection 2017. Front Syst Neurosci. 2017. PMID: 28420968 Free PMC article. No abstract available.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
