

Genome-Wide Transcriptional Regulation Mediated by Biochemically Distinct SWI/SNF Complexes
- PMID: 26716708
- PMCID: PMC4699898
- DOI: 10.1371/journal.pgen.1005748
Genome-Wide Transcriptional Regulation Mediated by Biochemically Distinct SWI/SNF Complexes
Abstract
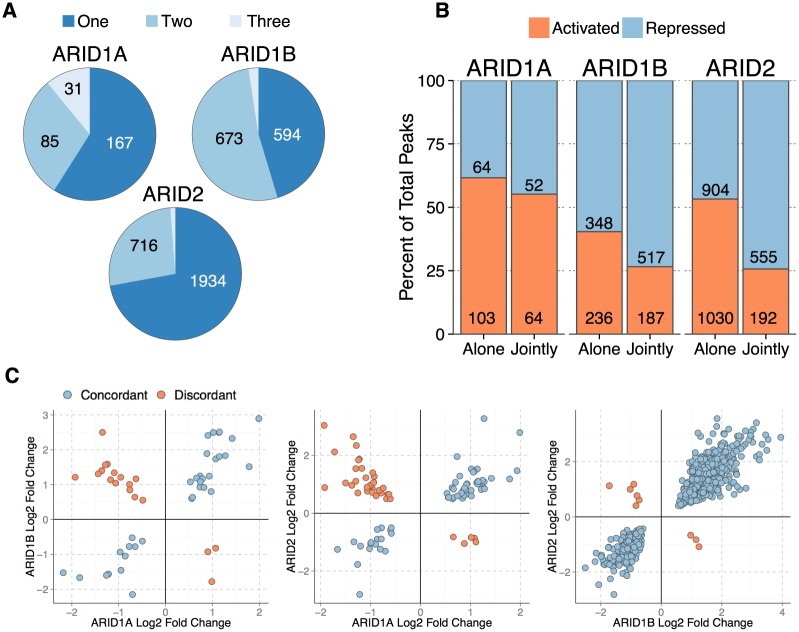
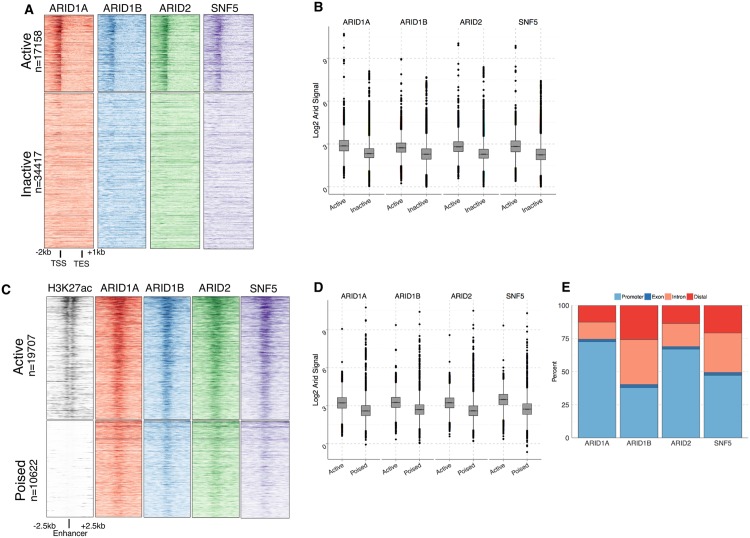
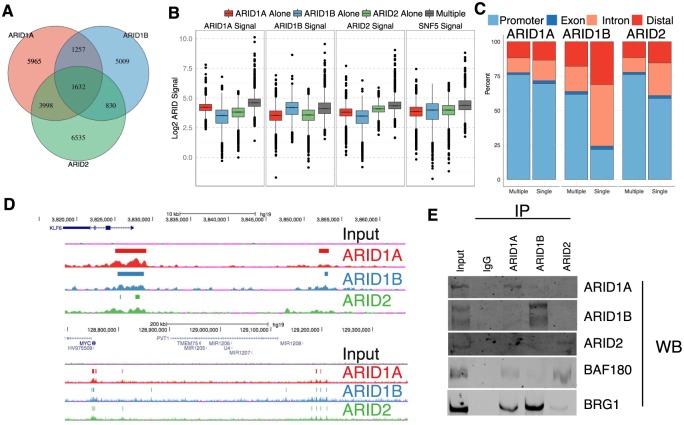
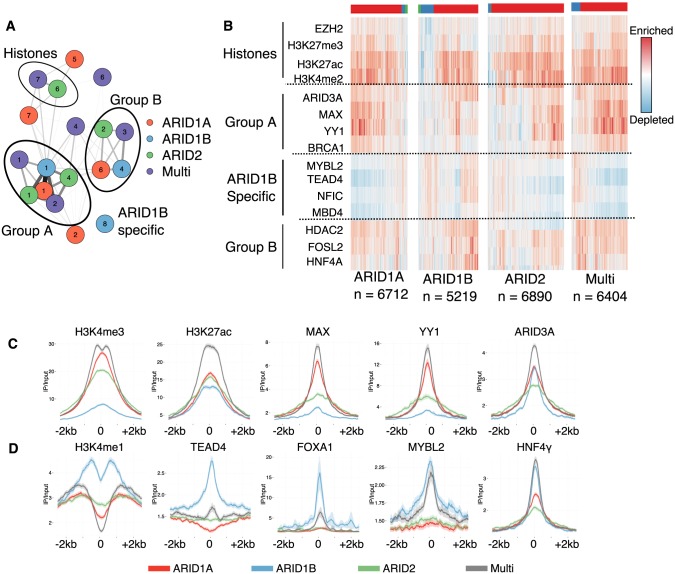
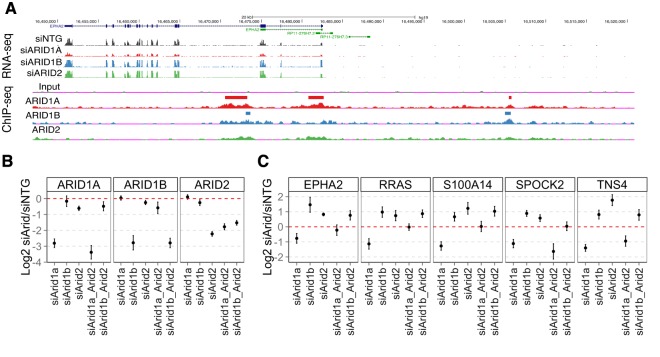
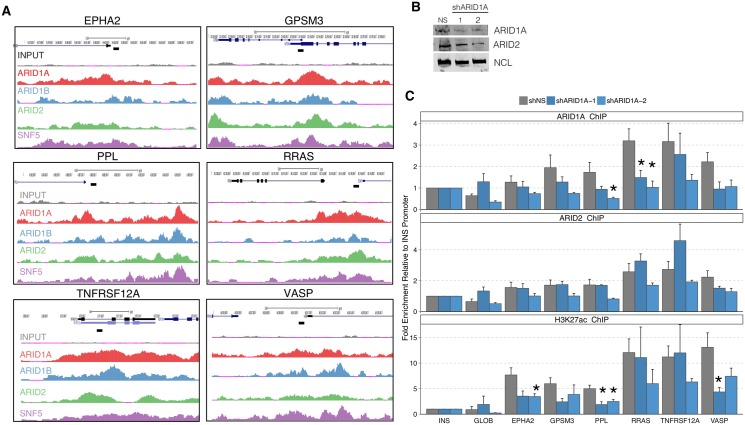
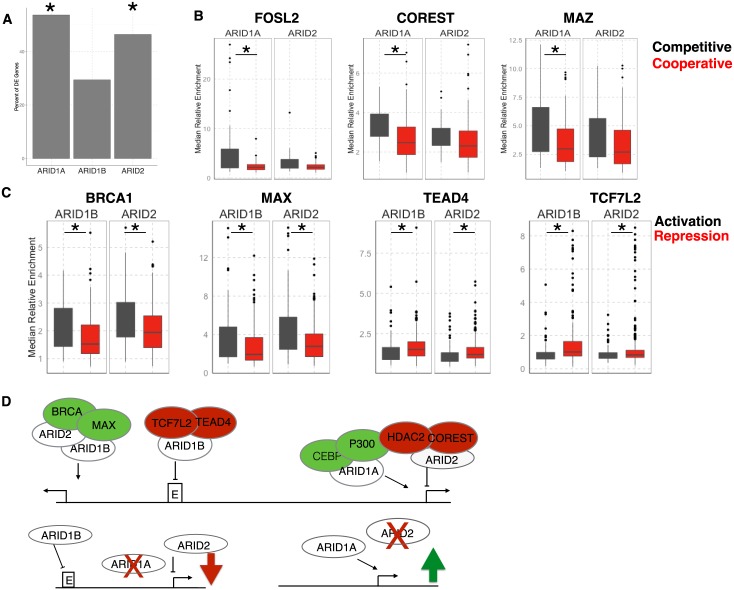
Multiple positions within the SWI/SNF chromatin remodeling complex can be filled by mutually exclusive subunits. Inclusion or exclusion of these proteins defines many unique forms of SWI/SNF and has profound functional consequences. Often this complex is studied as a single entity within a particular cell type and we understand little about the functional relationship between these biochemically distinct forms of the remodeling complex. Here we examine the functional relationships among three complex-specific ARID (AT-Rich Interacting Domain) subunits using genome-wide chromatin immunoprecipitation, transcriptome analysis, and transcription factor binding maps. We find widespread overlap in transcriptional regulation and the genomic binding of distinct SWI/SNF complexes. ARID1B and ARID2 participate in wide-spread cooperation to repress hundreds of genes. Additionally, we find numerous examples of competition between ARID1A and another ARID, and validate that gene expression changes following loss of one ARID are dependent on the function of an alternative ARID. These distinct regulatory modalities are correlated with differential occupancy by transcription factors. Together, these data suggest that distinct SWI/SNF complexes dictate gene-specific transcription through functional interactions between the different forms of the SWI/SNF complex and associated co-factors. Most genes regulated by SWI/SNF are controlled by multiple biochemically distinct forms of the complex, and the overall expression of a gene is the product of the interaction between these different SWI/SNF complexes. The three mutually exclusive ARID family members are among the most frequently mutated chromatin regulators in cancer, and understanding the functional interactions and their role in transcriptional regulation provides an important foundation to understand their role in cancer.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Ho L, Jothi R, Ronan JL, Cui K, Zhao K, Crabtree GR. An embryonic stem cell chromatin remodeling complex esBAF is an essential component of the core pluripotency transcriptional network. Proceedings of the National Academy of Sciences. 2009. March;106(13):5187–5191. Available from: 10.1073/pnas.0812888106 10.1073/pnas.0812889106 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
