

Detection of Quiescent Infections with Multiple Elephant Endotheliotropic Herpesviruses (EEHVs), Including EEHV2, EEHV3, EEHV6, and EEHV7, within Lymphoid Lung Nodules or Lung and Spleen Tissue Samples from Five Asymptomatic Adult African Elephants
- PMID: 26719245
- PMCID: PMC4810643
- DOI: 10.1128/JVI.02936-15
Detection of Quiescent Infections with Multiple Elephant Endotheliotropic Herpesviruses (EEHVs), Including EEHV2, EEHV3, EEHV6, and EEHV7, within Lymphoid Lung Nodules or Lung and Spleen Tissue Samples from Five Asymptomatic Adult African Elephants
Abstract
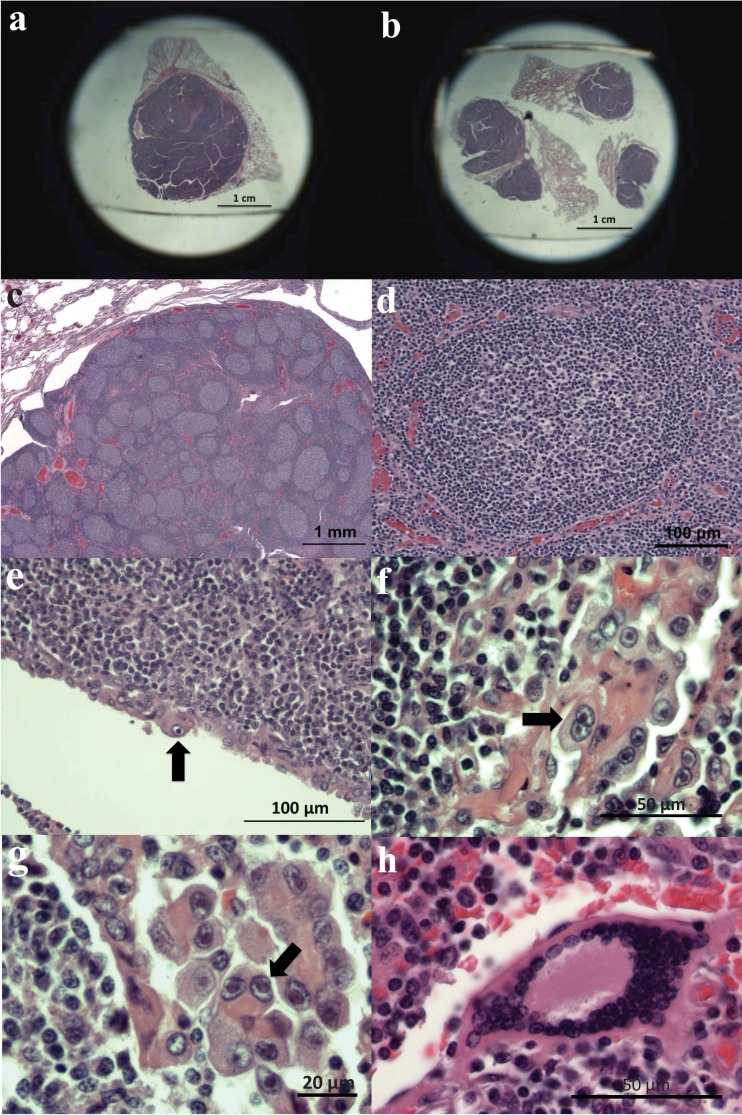
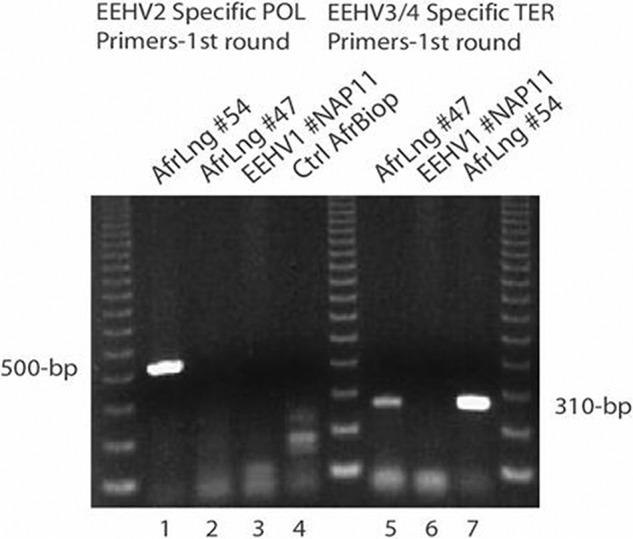
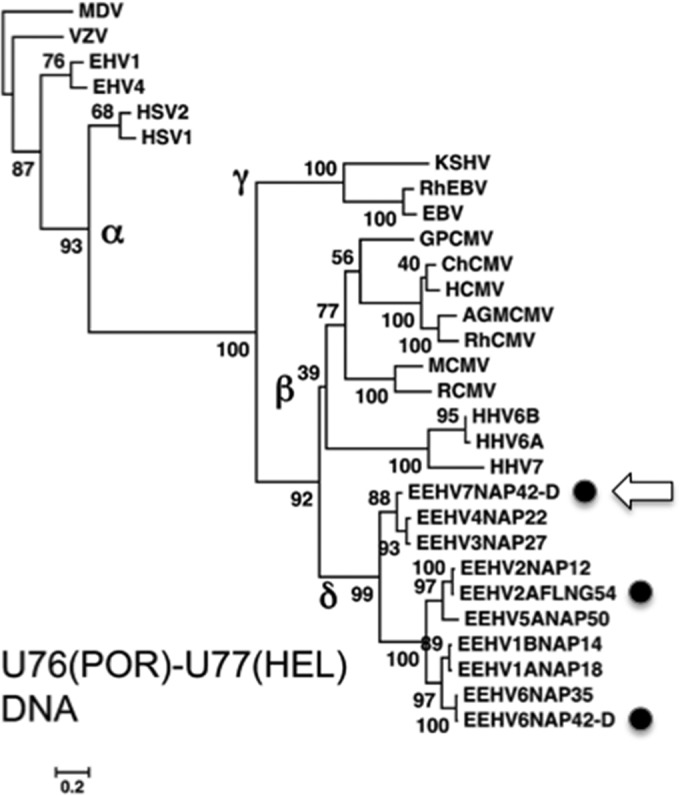
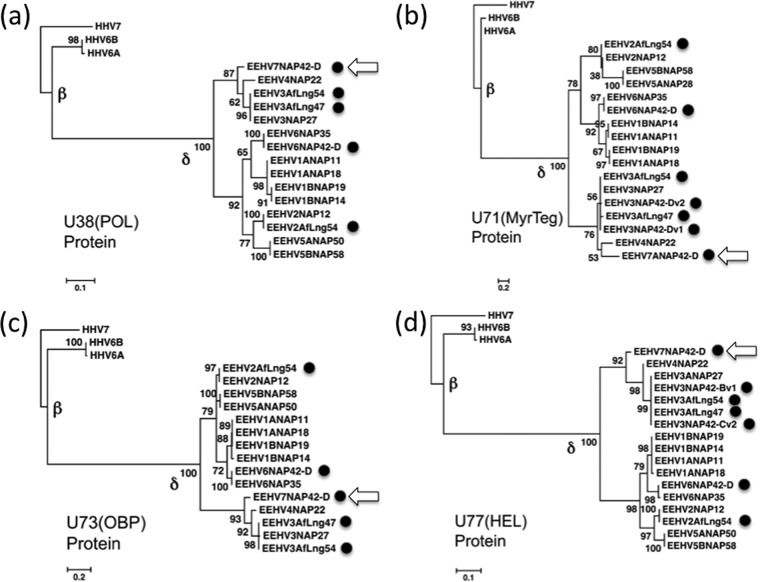
More than 80 cases of lethal hemorrhagic disease associated with elephant endotheliotropic herpesviruses (EEHVs) have been identified in young Asian elephants worldwide. Diagnostic PCR tests detected six types of EEHV in blood of elephants with acute disease, although EEHV1A is the predominant pathogenic type. Previously, the presence of herpesvirus virions within benign lung and skin nodules from healthy African elephants led to suggestions that African elephants may be the source of EEHV disease in Asian elephants. Here, we used direct PCR-based DNA sequencing to detect EEHV genomes in necropsy tissue from five healthy adult African elephants. Two large lung nodules collected from culled wild South African elephants contained high levels of either EEHV3 alone or both EEHV2 and EEHV3. Similarly, a euthanized U.S. elephant proved to harbor multiple EEHV types distributed nonuniformly across four small lung nodules, including high levels of EEHV6, lower levels of EEHV3 and EEHV2, and a new GC-rich branch type, EEHV7. Several of the same EEHV types were also detected in random lung and spleen samples from two other elephants. Sanger PCR DNA sequence data comprising 100 kb were obtained from a total of 15 different strains identified, with (except for a few hypervariable genes) the EEHV2, EEHV3, and EEHV6 strains all being closely related to known genotypes from cases of acute disease, whereas the seven loci (4.0 kb) obtained from EEHV7 averaged 18% divergence from their nearest relative, EEHV3. Overall, we conclude that these four EEHV species, but probably not EEHV1, occur commonly as quiescent infections in African elephants.
Importance: Acute hemorrhagic disease characterized by high-level viremia due to infection by members of the Proboscivirus genus threatens the future breeding success of endangered Asian elephants worldwide. Although the genomes of six EEHV types from acute cases have been partially or fully characterized, lethal disease predominantly involves a variety of strains of EEHV1, whose natural host has been unclear. Here, we carried out genotype analyses by partial PCR sequencing of necropsy tissue from five asymptomatic African elephants and identified multiple simultaneous infections by several different EEHV types, including high concentrations in lymphoid lung nodules. Overall, the results provide strong evidence that EEHV2, EEHV3, EEHV6, and EEHV7 represent natural ubiquitous infections in African elephants, whereas Asian elephants harbor EEHV1A, EEHV1B, EEHV4, and EEHV5. Although a single case of fatal cross-species infection by EEHV3 is known, the results do not support the previous concept that highly pathogenic EEHV1A crossed from African to Asian elephants in zoos.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Richman LK, Montali RJ, Garber RL, Kennedy MA, Lehnhardt J, Hildebrandt T, Schmitt D, Hardy D, Alcendor DJ, Hayward GS. 1999. Novel endotheliotropic herpesviruses fatal for Asian and African elephants. Science 283:1171–1176. - PubMed
-
- Latimer E, Zong J-C, Heaggans SY, Richman LK, Hayward GS. 2011. Detection and evaluation of novel herpesviruses in routine and pathological samples from Asian and African elephants: identification of two new probosciviruses (EEHV5 and EEHV6) and two new gammaherpesviruses (EGHV3B and EGHV5). Vet Microbiol 147:28–41. doi:10.1016/j.vetmic.2010.05.042. - DOI - PMC - PubMed
-
- Richman LK, Zong J-C, Latimer E, Lock J, Fleischer RC, Heaggans SY, Hayward GS. 2014. Elephant endotheliotropic herpesviruses EEHV1A, EEHV1B, and EEHV2 from cases of hemorrhagic disease are highly diverged from other mammalian herpesviruses and may form a new subfamily. J Virol 88:13523–13546. doi:10.1128/JVI.01673-14. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
