

Human Cytomegalovirus US28 Is Important for Latent Infection of Hematopoietic Progenitor Cells
- PMID: 26719258
- PMCID: PMC4810657
- DOI: 10.1128/JVI.02507-15
Human Cytomegalovirus US28 Is Important for Latent Infection of Hematopoietic Progenitor Cells
Abstract
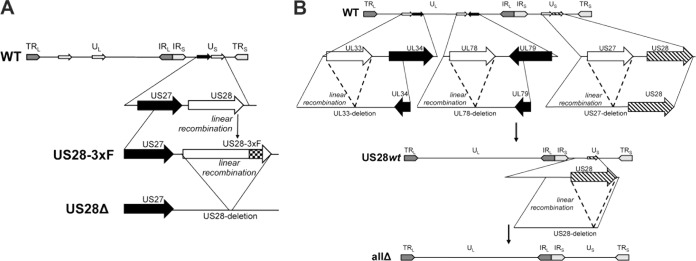
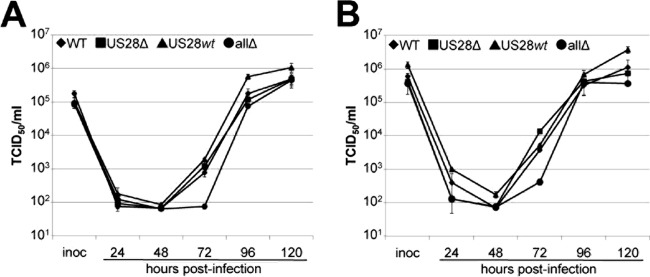
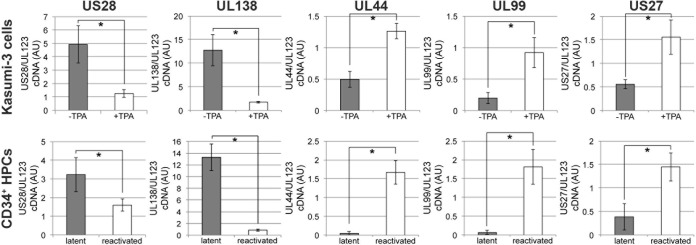
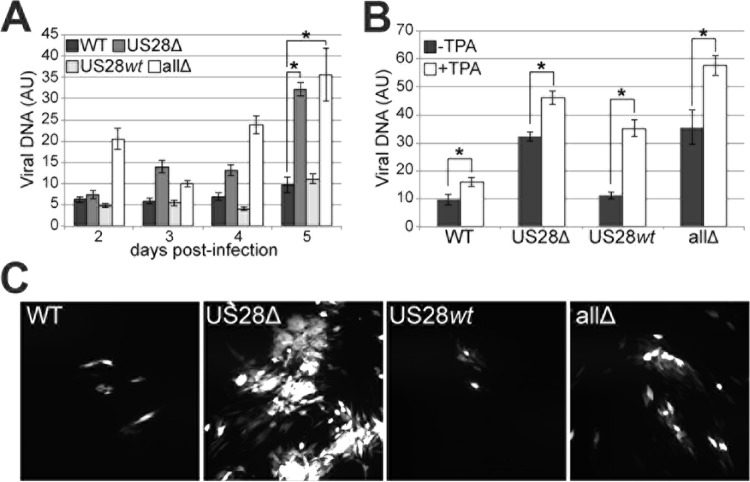
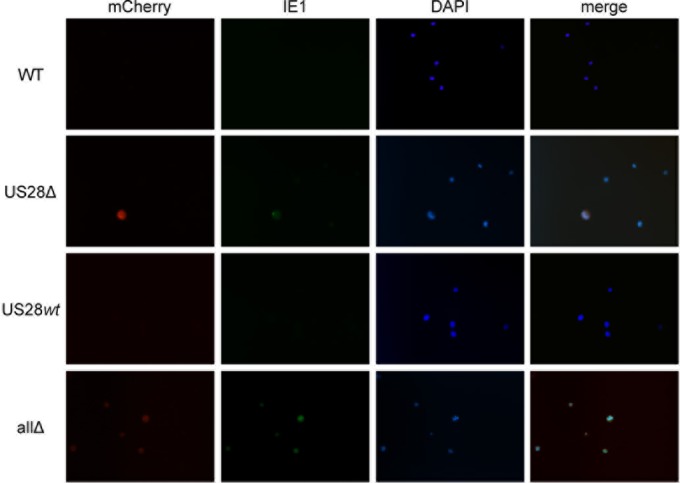
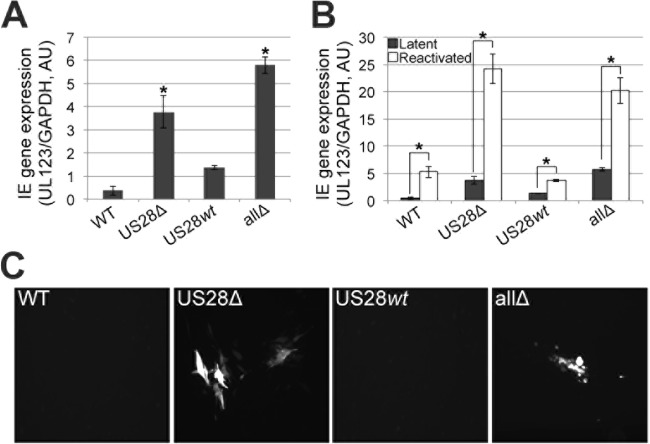
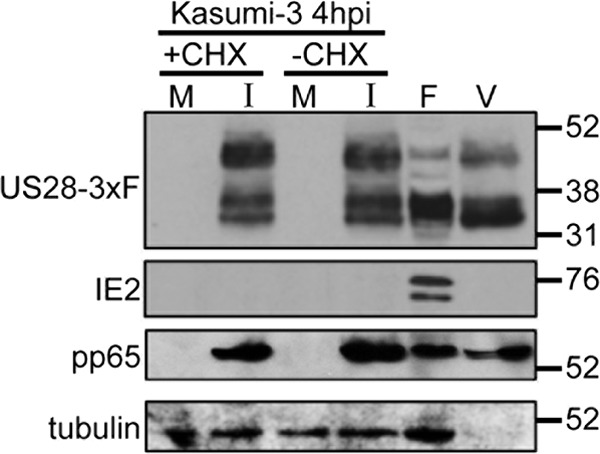
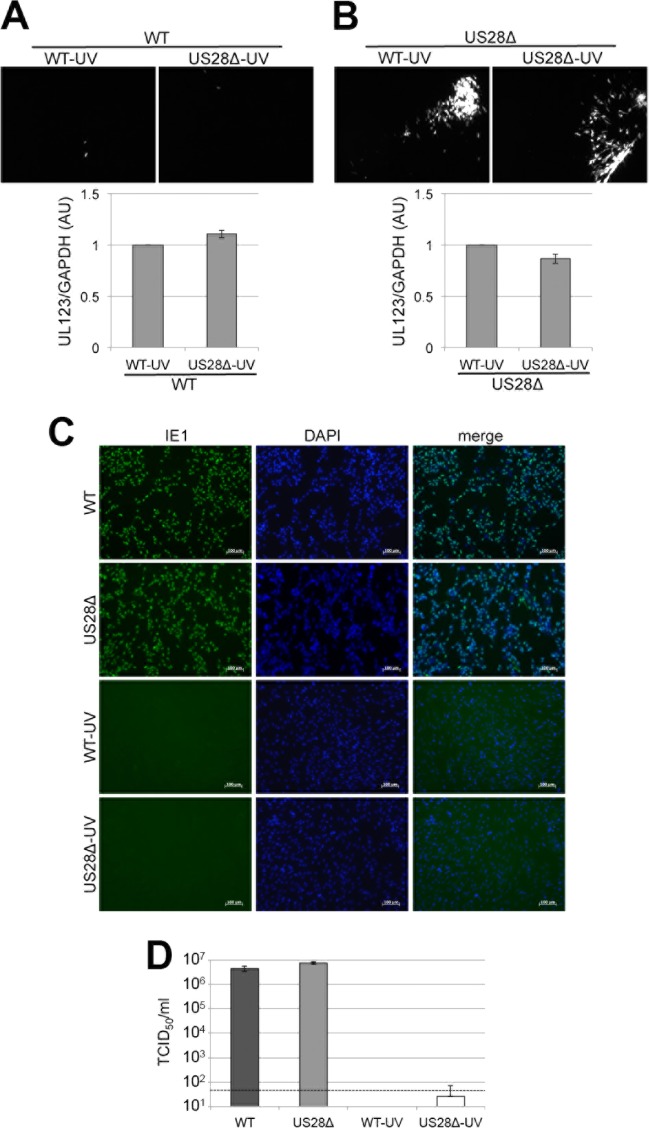
Human cytomegalovirus (HCMV) resides latently in hematopoietic progenitor cells (HPCs). During latency, only a subset of HCMV genes is transcribed, including one of the four virus-encoded G protein-coupled receptors (GPCRs), US28. Although US28 is a multifunctional lytic protein, its function during latency has remained undefined. We generated a panel of US28 recombinant viruses in the bacterial artificial chromosome (BAC)-derived clinical HCMV strain TB40/E-mCherry. We deleted the entire US28 open reading frame (ORF), deleted all four of the viral GPCR ORFs, or deleted three of the HCMV GPCRs but not the US28 wild-type protein. Using these recombinant viruses, we assessed the requirement for US28 during latency in the Kasumi-3 in vitro latency model system and in primary ex vivo-cultured CD34(+) HPCs. Our data suggest that US28 is required for latency as infection with viruses lacking the US28 ORF alone or in combination with the remaining HCMV-encoded GPCR results in transcription from the major immediate early promoter, the production of extracellular virions, and the production of infectious virus capable of infecting naive fibroblasts. The other HCMV GPCRs are not required for this phenotype as a virus expressing only US28 but not the remaining virus-encoded GPCRs is phenotypically similar to that of wild-type latent infection. Finally, we found that US28 copurifies with mature virions and is expressed in HPCs upon virus entry although its expression at the time of infection does not complement the US28 deletion latency phenotype. This work suggests that US28 protein functions to promote a latent state within hematopoietic progenitor cells.
Importance: Human cytomegalovirus (HCMV) is a widespread pathogen that, once acquired, remains with its host for life. HCMV remains latent, or quiescent, in cells of the hematopoietic compartment and upon immune challenge can reactivate to cause disease. HCMV-encoded US28 is one of several genes expressed during latency although its biological function during this phase of infection has remained undefined. Here, we show that US28 aids in promoting experimental latency in tissue culture.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Britt W. 2008. Manifestations of human cytomegalovirus infection: proposed mechanisms of acute and chronic disease. Curr Top Microbiol Immunol 325:417–470. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
