

Myocyte-derived Tnfsf14 is a survival factor necessary for myoblast differentiation and skeletal muscle regeneration
- PMID: 26720335
- PMCID: PMC4720906
- DOI: 10.1038/cddis.2015.375
Myocyte-derived Tnfsf14 is a survival factor necessary for myoblast differentiation and skeletal muscle regeneration
Abstract
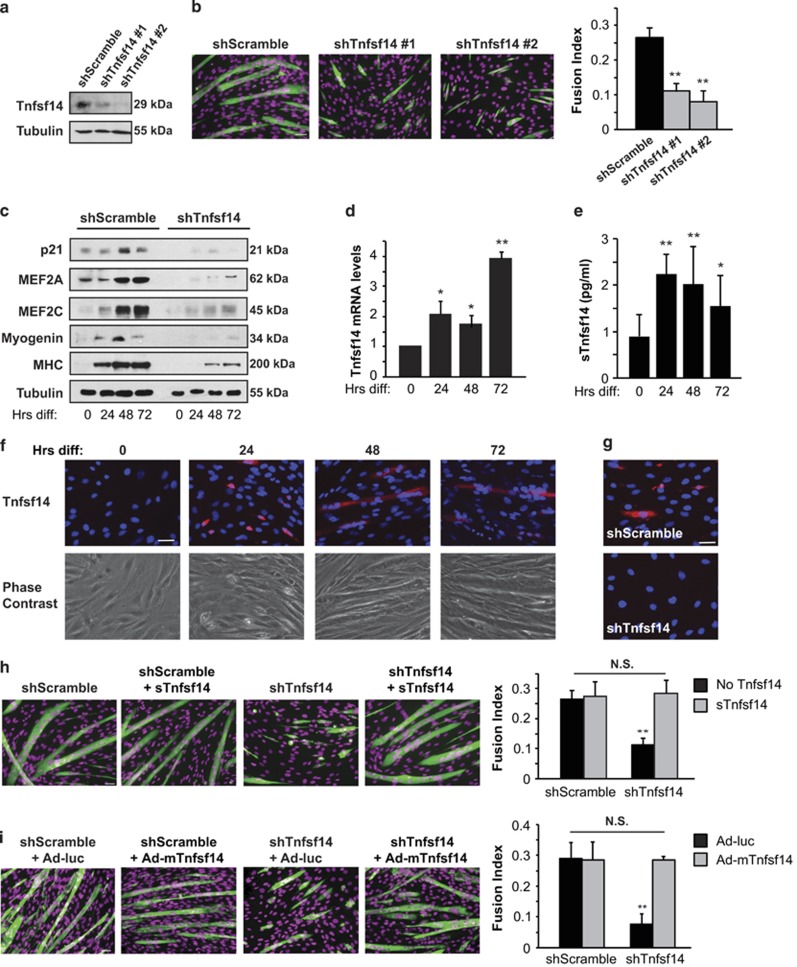
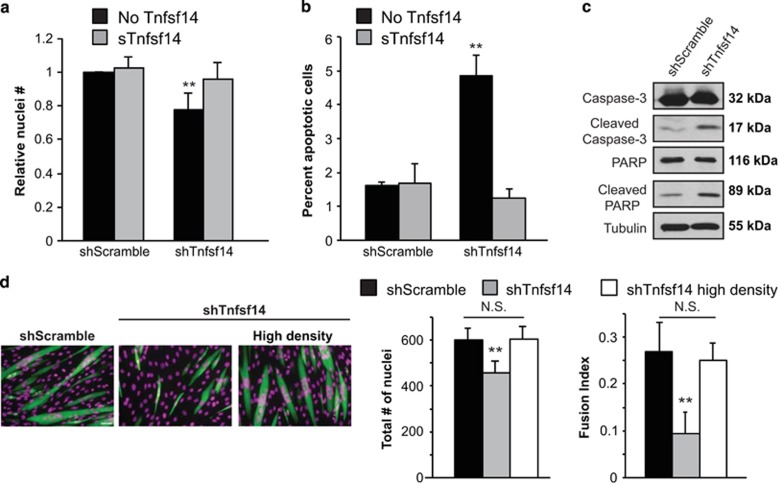
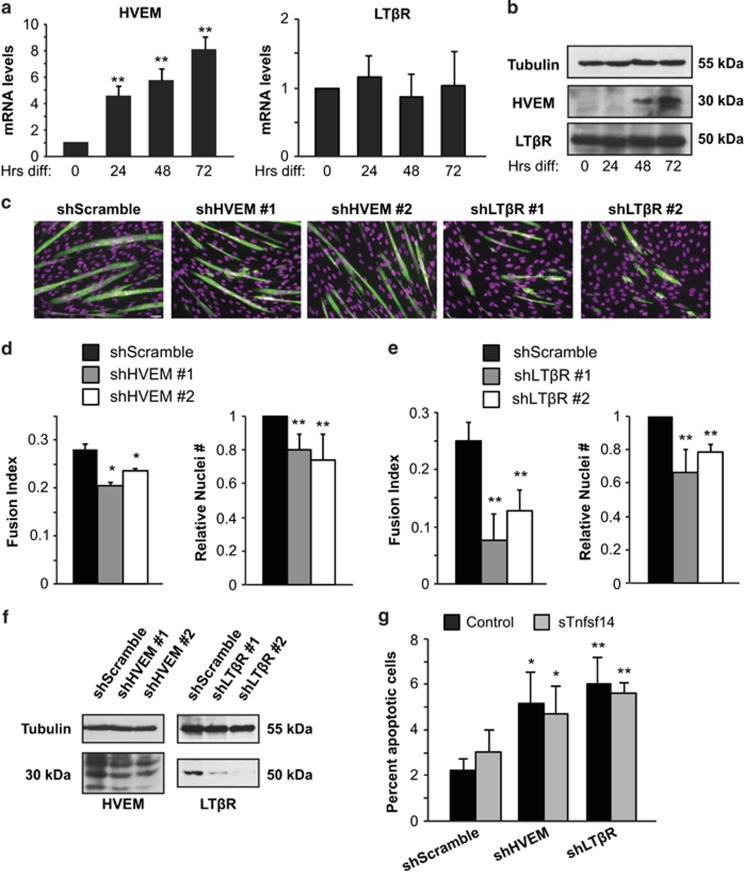
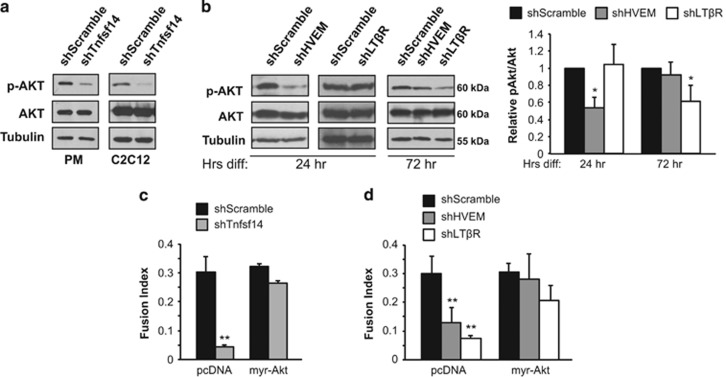
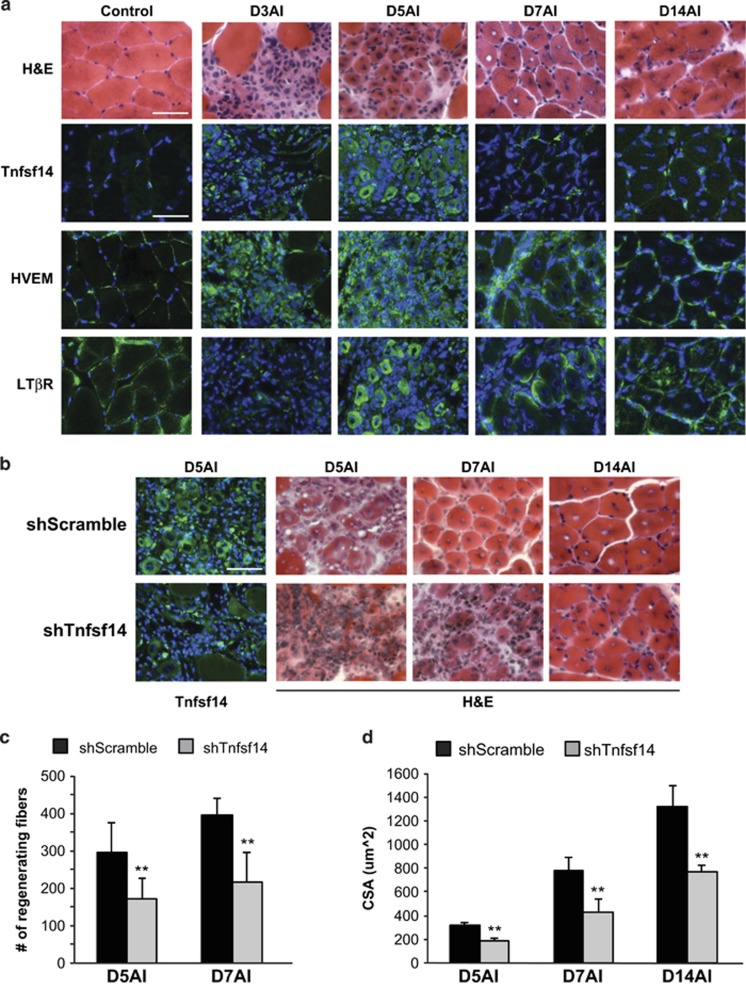
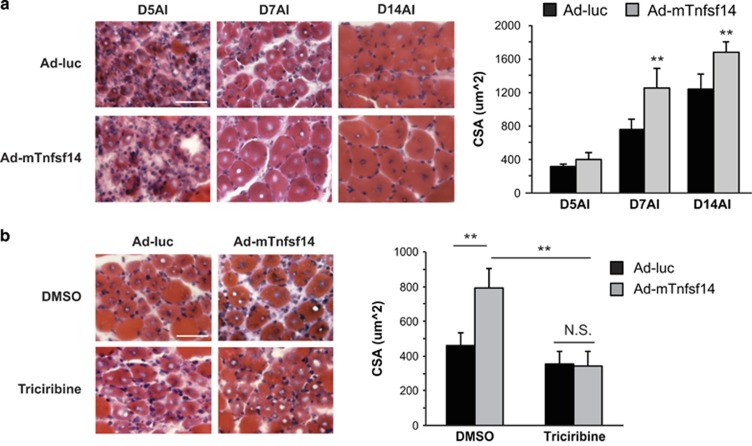
Adult skeletal muscle tissue has a uniquely robust capacity for regeneration, which gradually declines with aging or is compromised in muscle diseases. The cellular mechanisms regulating adult myogenesis remain incompletely understood. Here we identify the cytokine tumor necrosis factor superfamily member 14 (Tnfsf14) as a positive regulator of myoblast differentiation in culture and muscle regeneration in vivo. We find that Tnfsf14, as well as its cognate receptors herpes virus entry mediator (HVEM) and lymphotoxin β receptor (LTβR), are expressed in both differentiating myocytes and regenerating myofibers. Depletion of Tnfsf14 or either receptor inhibits myoblast differentiation and promotes apoptosis. Our results also suggest that Tnfsf14 regulates myogenesis by supporting cell survival and maintaining a sufficient pool of cells for fusion. In addition, we show that Akt mediates the survival and myogenic function of Tnfsf14. Importantly, local knockdown of Tnfsf14 is found to impair injury-induced muscle regeneration in a mouse model, affirming an important physiological role for Tnfsf14 in myogenesis in vivo. Furthermore, we demonstrate that localized overexpression of Tnfsf14 potently enhances muscle regeneration, and that this regenerative capacity of Tnfsf14 is dependent on Akt signaling. Taken together, our findings reveal a novel regulator of skeletal myogenesis and implicate Tnfsf14 in future therapeutic development.
Figures
References
-
- 1Relaix F, Marcelle C. Muscle stem cells. Curr Opin Cell Biol 2009; 21: 748. - PubMed
-
- 2Tidball JG. Mechanisms of muscle injury, repair, and regeneration. Compr Physiol 2011; 1: 2029. - PubMed
-
- 3Naya FS, Olson E. MEF2: a transcriptional target for signaling pathways controlling skeletal muscle growth and differentiation. Curr Opin Cell Biol 1999; 11: 683. - PubMed
-
- 4Parker MH, Seale P, Rudnicki MA. Looking back to the embryo: defining transcriptional networks in adult myogenesis. Nat Rev Genet 2003; 4: 497. - PubMed
-
- 5Wagers AJ, Conboy IM. Cellular and molecular signatures of muscle regeneration: current concepts and controversies in adult myogenesis. Cell 2005; 122: 659. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
