

The basal function of teleost prolactin as a key regulator on ion uptake identified with zebrafish knockout models
- PMID: 26726070
- PMCID: PMC4698586
- DOI: 10.1038/srep18597
The basal function of teleost prolactin as a key regulator on ion uptake identified with zebrafish knockout models
Abstract
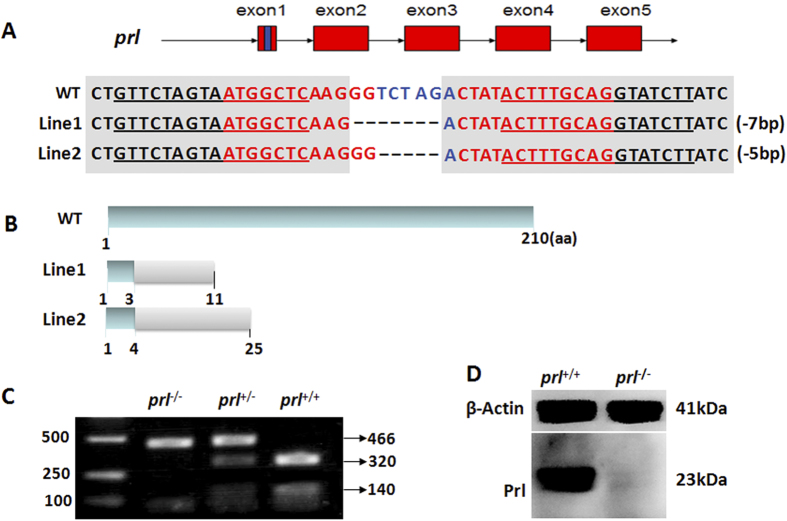
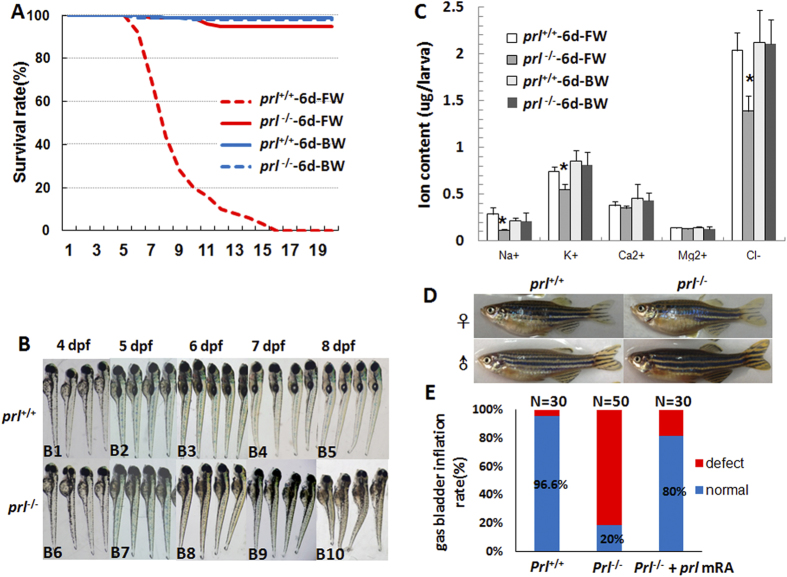
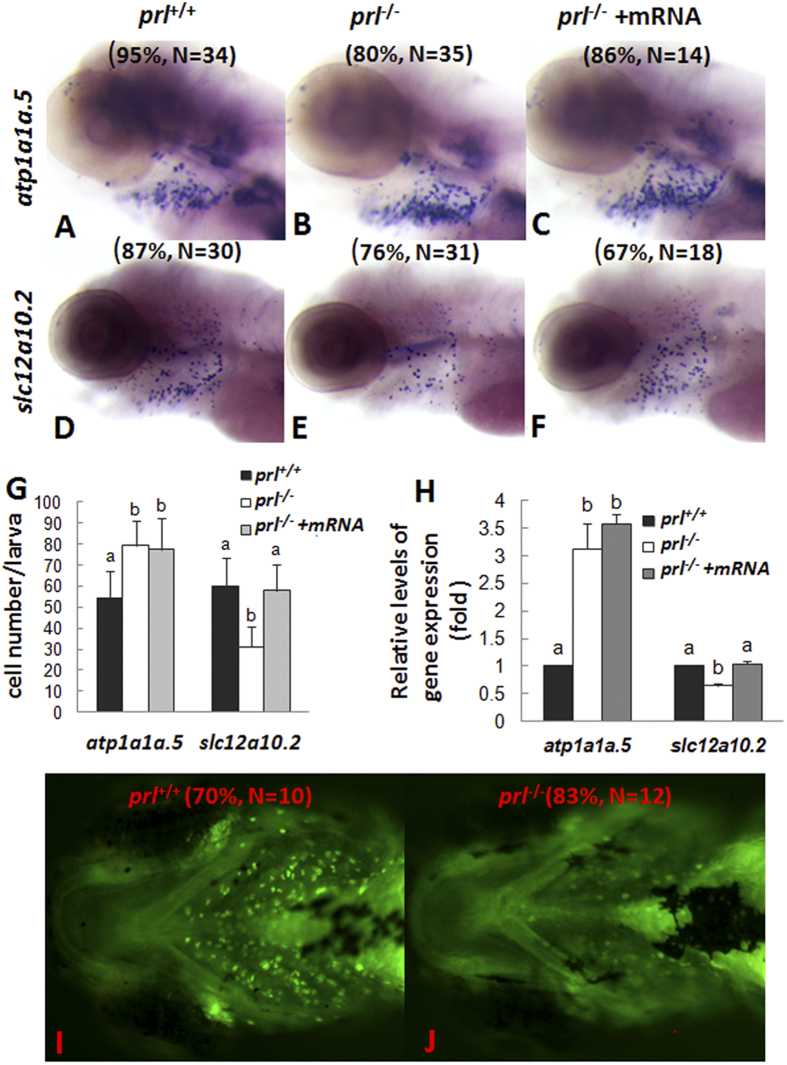
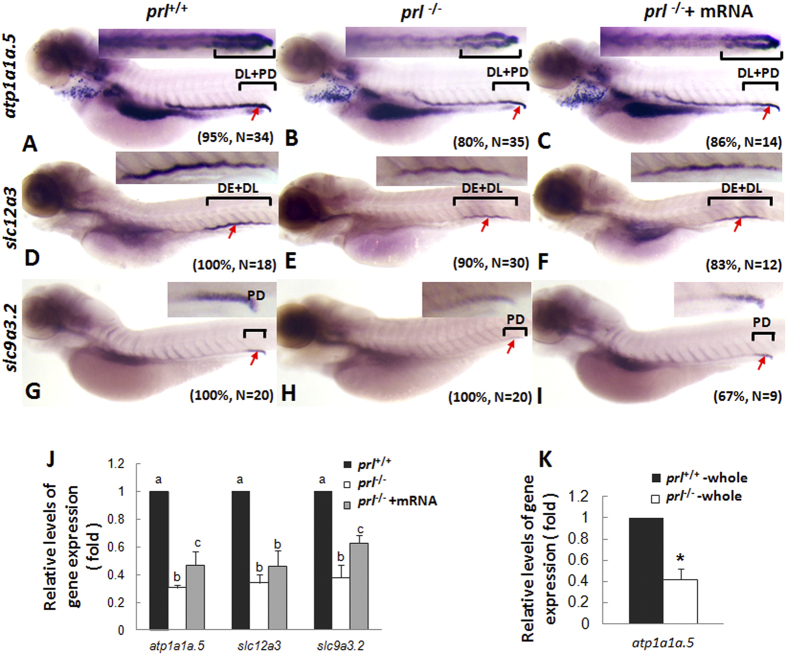
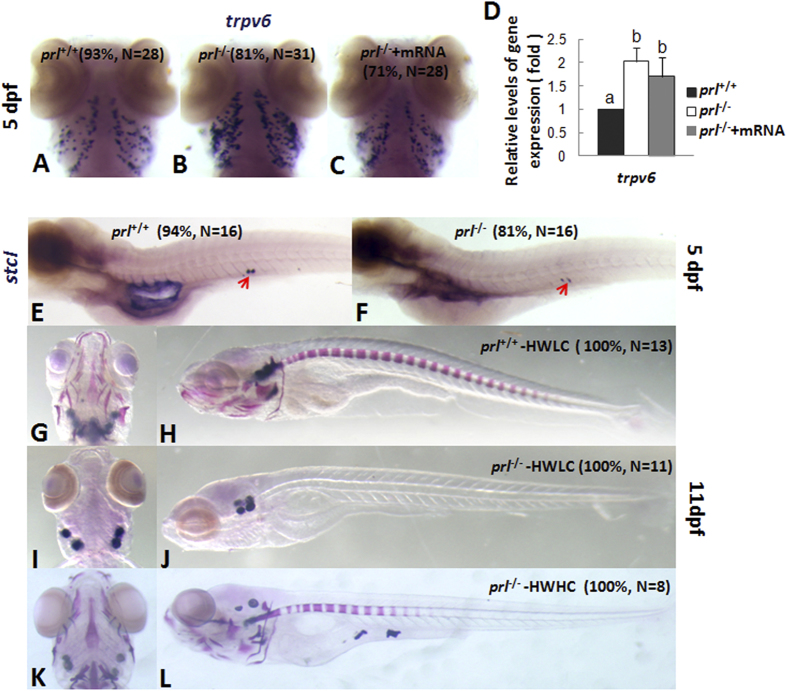
Prolactin (PRL) is an anterior pituitary hormone with a broad range of functions. Its ability to stimulate lactogenesis, maternal behavior, growth and development, osmoregulation, and epithelial ion transport has been reported in many vertebrates. In our present study, we have targeted the zebrafish prl locus via transcription activator-like effector nucleases (TALENs). Two independent targeted mutant lines with premature termination of the putative sequence of PRL peptides were generated. All prl-deficient zebrafish progeny died at 6-16 days post-fertilization stage (dpf) in egg water. However, the prl-deficient larvae thrived and survived through adulthood in brackish water (5175 mg/L ocean salts), without obvious defects in somatic growth or reproduction. When raised in egg water, the expression levels of certain key Na(+)/Cl(-) cotransporters in the gills and Na(+)/K(+)-ATPase subunits, Na(+)/H(+) exchangers and Na(+)/Cl(-) transporters in the pronephros of prl-deficient larvae were down-regulated at 5 dpf, which caused Na(+)/K(+)/Cl(-) uptake defects in the mutant fish at 6 dpf. Our present results demonstrate that the primary function of zebrafish prl is osmoregulation via governing the uptake and homeostasis of Na(+), K(+) and Cl(-). Our study provides valuable evidence to understand the mechanisms of PRL function better through both phylogenetic and physiological perspectives.
Figures

Similar articles
-
Depletion of Tissue-Specific Ion Transporters Causes Differential Expression of PRL Targets in Response to Increased Levels of Endogenous PRL.Front Endocrinol (Lausanne). 2018 Nov 20;9:683. doi: 10.3389/fendo.2018.00683. eCollection 2018. Front Endocrinol (Lausanne). 2018. PMID: 30515132 Free PMC article.
-
Prolactin regulates transcription of the ion uptake Na+/Cl- cotransporter (ncc) gene in zebrafish gill.Mol Cell Endocrinol. 2013 Apr 30;369(1-2):98-106. doi: 10.1016/j.mce.2013.01.021. Epub 2013 Feb 6. Mol Cell Endocrinol. 2013. PMID: 23395804 Free PMC article.
-
The involvement of SLC26 anion transporters in chloride uptake in zebrafish (Danio rerio) larvae.J Exp Biol. 2009 Oct;212(Pt 20):3283-95. doi: 10.1242/jeb.033910. J Exp Biol. 2009. PMID: 19801433
-
Prolactin-dependent modulation of organogenesis in the vertebrate: Recent discoveries in zebrafish.Comp Biochem Physiol C Toxicol Pharmacol. 2008 Nov;148(4):370-80. doi: 10.1016/j.cbpc.2008.05.010. Epub 2008 May 28. Comp Biochem Physiol C Toxicol Pharmacol. 2008. PMID: 18593647 Review.
-
The role of prolactin in fish osmoregulation: a review.Gen Comp Endocrinol. 2002 Feb 1;125(2):291-310. doi: 10.1006/gcen.2001.7746. Gen Comp Endocrinol. 2002. PMID: 11884075 Review.
Cited by
-
Secretion and Function of Pituitary Prolactin in Evolutionary Perspective.Front Neurosci. 2020 Jun 16;14:621. doi: 10.3389/fnins.2020.00621. eCollection 2020. Front Neurosci. 2020. PMID: 32612510 Free PMC article. Review.
-
Stanniocalcin 1a regulates organismal calcium balance and survival by suppressing Trpv6 expression and inhibiting IGF signaling in zebrafish.Front Endocrinol (Lausanne). 2023 Oct 26;14:1276348. doi: 10.3389/fendo.2023.1276348. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 37964974 Free PMC article.
-
Depletion of Tissue-Specific Ion Transporters Causes Differential Expression of PRL Targets in Response to Increased Levels of Endogenous PRL.Front Endocrinol (Lausanne). 2018 Nov 20;9:683. doi: 10.3389/fendo.2018.00683. eCollection 2018. Front Endocrinol (Lausanne). 2018. PMID: 30515132 Free PMC article.
-
Type II Na+-phosphate Cotransporters and Phosphate Balance in Teleost Fish.Pflugers Arch. 2019 Jan;471(1):193-212. doi: 10.1007/s00424-018-2239-4. Epub 2018 Dec 12. Pflugers Arch. 2019. PMID: 30542786 Review.
-
Heavy metal toxicity in Buriganga river alters the immunology of Nile tilapia (Oreochromis niloticus L).Heliyon. 2021 Oct 28;7(11):e08285. doi: 10.1016/j.heliyon.2021.e08285. eCollection 2021 Nov. Heliyon. 2021. PMID: 34765796 Free PMC article.
References
-
- Riddle O., Bates R. W. & Dykshorn S. W. The preparation, identification and assay of prolactin - A hormone of the anterior pituitary. Am J Physiol 105, 191–216 (1933).
-
- Ormandy C. J. et al. Null mutation of the prolactin receptor gene produces multiple reproductive defects in the mouse. Genes & development 11, 167–178 (1997). - PubMed
-
- Horseman N. D. & Gregerson K. A. Prolactin actions. J Mol Endocrinol 52, R95–106 (2014). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
