

LmABCB3, an atypical mitochondrial ABC transporter essential for Leishmania major virulence, acts in heme and cytosolic iron/sulfur clusters biogenesis
- PMID: 26728034
- PMCID: PMC4700571
- DOI: 10.1186/s13071-015-1284-5
LmABCB3, an atypical mitochondrial ABC transporter essential for Leishmania major virulence, acts in heme and cytosolic iron/sulfur clusters biogenesis
Abstract
Background: Mitochondria play essential biological functions including the synthesis and trafficking of porphyrins and iron/sulfur clusters (ISC), processes that in mammals involve the mitochondrial ATP-Binding Cassette (ABC) transporters ABCB6 and ABCB7, respectively. The mitochondrion of pathogenic protozoan parasites such as Leishmania is a promising goal for new therapeutic approaches. Leishmania infects human macrophages producing the neglected tropical disease known as leishmaniasis. Like most trypanosomatid parasites, Leishmania is auxotrophous for heme and must acquire porphyrins from the host.
Methods: LmABCB3, a new Leishmania major protein with significant sequence similarity to human ABCB6/ABCB7, was identified and characterized using bioinformatic tools. Fluorescent microscopy was used to determine its cellular localization, and its level of expression was modulated by molecular genetic techniques. Intracellular in vitro assays were used to demonstrate its role in amastigotes replication, and an in vivo mouse model was used to analyze its role in virulence. Functional characterization of LmABCB3 was carried out in Leishmania promastigotes and Saccharomyces cerevisiae. Structural analysis of LmABCB3 was performed using molecular modeling software.
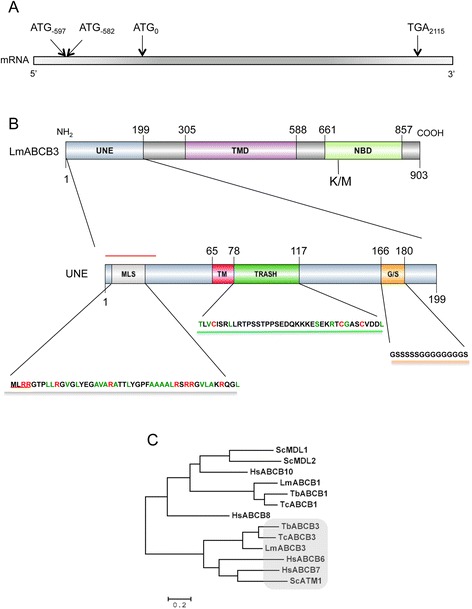
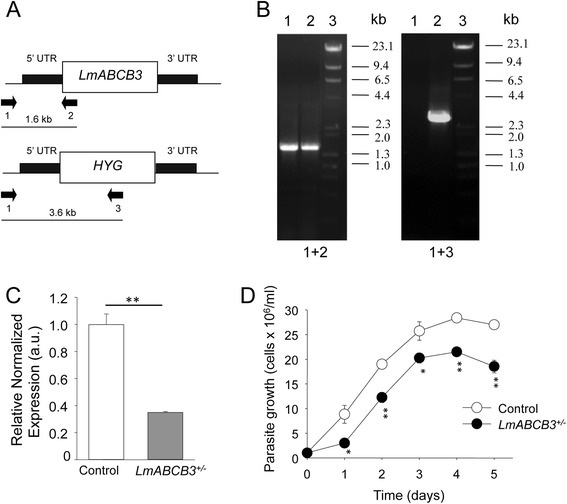
Results: LmABCB3 is an atypical ABC half-transporter that has a unique N-terminal extension not found in any other known ABC protein. This extension is required to target LmABCB3 to the mitochondrion and includes a potential metal-binding domain. We have shown that LmABCB3 interacts with porphyrins and is required for the mitochondrial synthesis of heme from a host precursor. We also present data supporting a role for LmABCB3 in the biogenesis of cytosolic ISC, essential cofactors for cell viability in all three kingdoms of life. LmABCB3 fully complemented the severe growth defect shown in yeast lacking ATM1, an orthologue of human ABCB7 involved in exporting from the mitochondria a gluthatione-containing compound required for the generation of cytosolic ISC. Indeed, docking analyzes performed with a LmABCB3 structural model using trypanothione, the main thiol in this parasite, as a ligand showed how both, LmABCB3 and yeast ATM1, contain a similar thiol-binding pocket. Additionally, we show solid evidence suggesting that LmABCB3 is an essential gene as dominant negative inhibition of LmABCB3 is lethal for the parasite. Moreover, the abrogation of only one allele of the gene did not impede promastigote growth in axenic culture but prevented the replication of intracellular amastigotes and the virulence of the parasites in a mouse model of cutaneous leishmaniasis.
Conclusions: Altogether our results present the previously undescribed LmABCB3 as an unusual mitochondrial ABC transporter essential for Leishmania survival through its role in the generation of heme and cytosolic ISC. Hence, LmABCB3 could represent a novel target to combat leishmaniasis.
Figures






References
-
- Trager W. Nutrition and biosynthetic capabilities of flagellates. Problems of in vitro cultivation and differentiation. In: CIBAFoundationSymposium, editor. Trypanosomiasis and Leishmaniasis with special reference to Chagas’ disease. North Holland, Amsterdam: Elsevier-Excerpta; 1974. pp. 225–245.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
