

Review
doi: 10.1083/jcb.201510032.
Defective control of pre-messenger RNA splicing in human disease
Affiliations
- PMID: 26728853
- PMCID: PMC4700483
- DOI: 10.1083/jcb.201510032
Item in Clipboard
Review
Defective control of pre-messenger RNA splicing in human disease
J Cell Biol.
.
Abstract
Examples of associations between human disease and defects in pre-messenger RNA splicing/alternative splicing are accumulating. Although many alterations are caused by mutations in splicing signals or regulatory sequence elements, recent studies have noted the disruptive impact of mutated generic spliceosome components and splicing regulatory proteins. This review highlights recent progress in our understanding of how the altered splicing function of RNA-binding proteins contributes to myelodysplastic syndromes, cancer, and neuropathologies.
© 2016 Chabot and Shkreta.
Figures
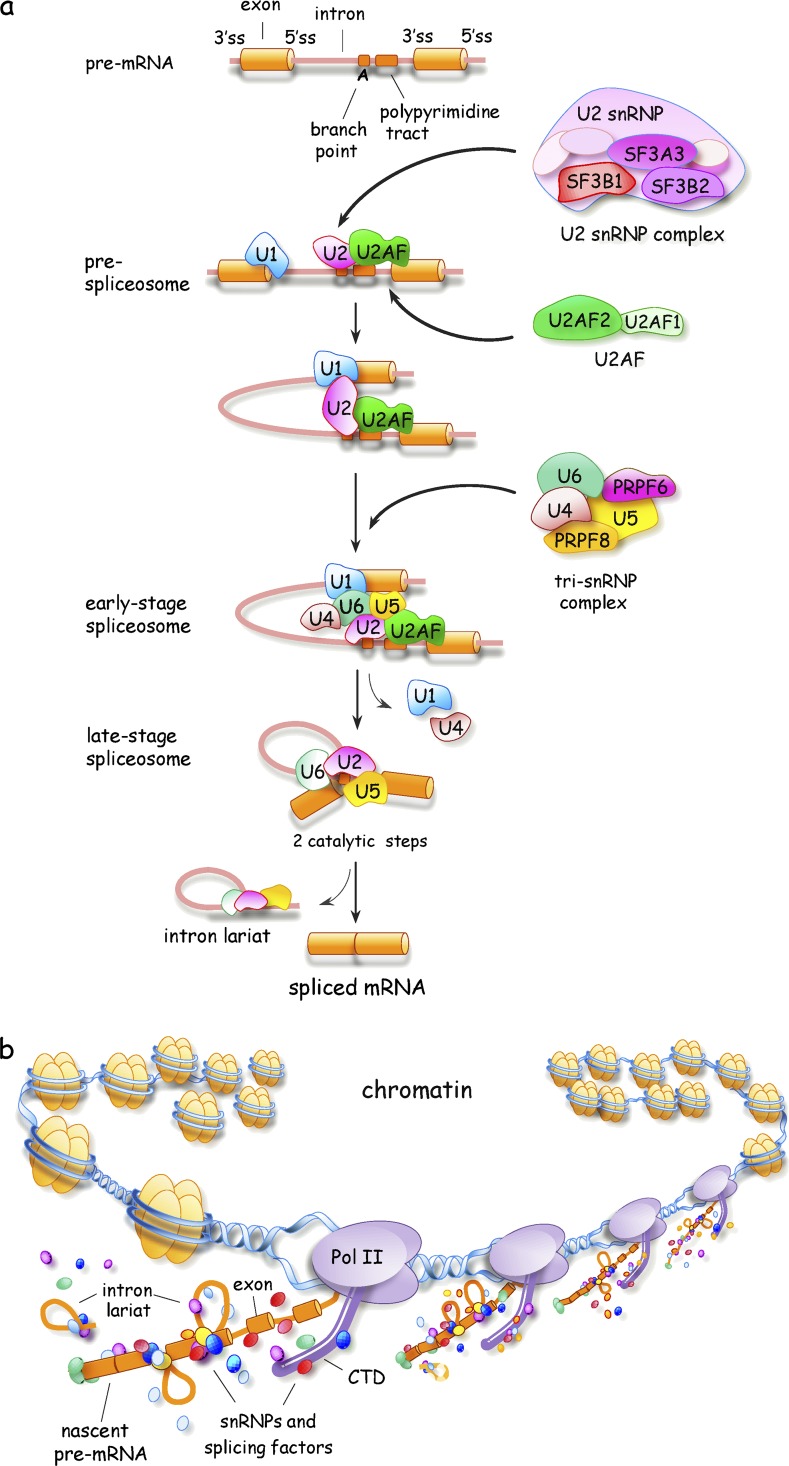
Spliceosome assembly and transcription-coupled splicing. (A) Schematic representation of spliceosome assembly indicating the position of 5′ss, 3′ss, the branch point, and the polypyrimidine tract. Exons and introns are represented as solid cylinders and lines, respectively. Only a portion of spliceosome components are depicted, with some subunits of U2AF, U2 snRNP, and the tri-snRNP complex indicated. (B) Schematic representation of the chromatin-associated cotranscriptional assembly of splicing complexes on a nascent pre-mRNA. CTD, C-terminal domain of RNA polymerase II.

Diagram summarizing how splicing factors and telomere deficiencies reproduce hematopoietic defects found in MDS. The impact of each deficiency on alternative splicing is indicated with an emphasis on shared alterations (see Spliceosomal proteins in MDS for details).
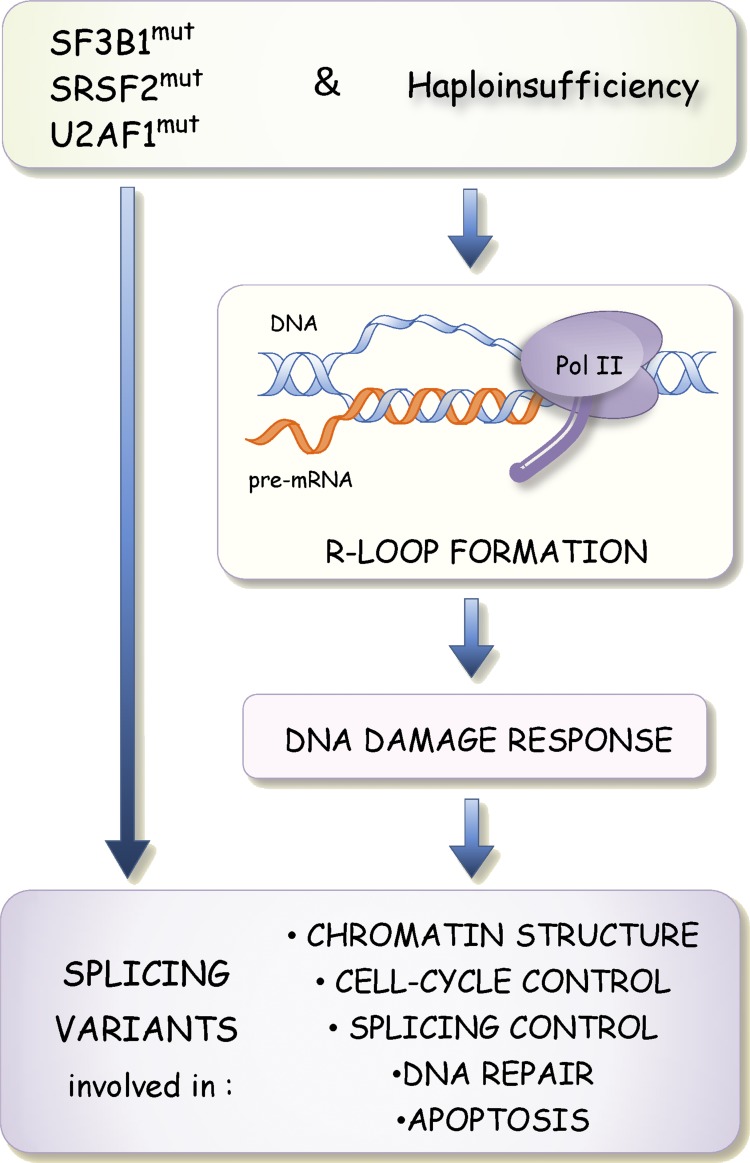
Model proposing that changes in alternative splicing triggered by mutations in SF3B1, SRSF2, and U2AF1 also occur through activation of the DDR. Drops in the activity or changes in the binding specificities of splicing factors alter splice site selection directly but are also proposed to promote the formation of R loops when segments of nascent pre-mRNAs are not adequately bound by splicing factors. R loop formation in turn triggers the DDR to elicit splicing changes in transcripts encoding DNA repair, chromatin structure, cell cycle and splicing control, and apoptotic components.
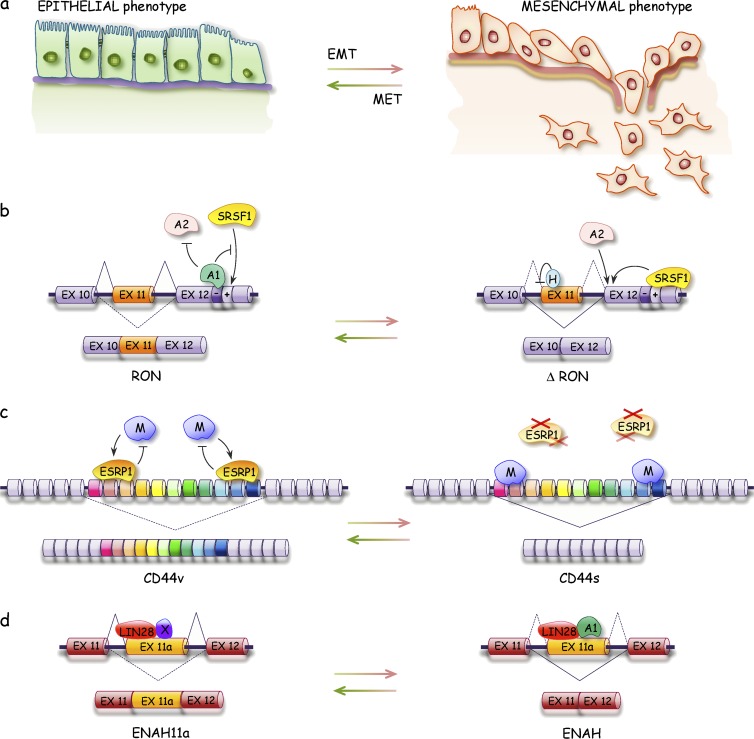
Defects in alternative splicing affect EMTs. (A–D) Epithelial and mesenchymal interconversion (A) are affected by splicing factors that control the alternative splicing events associated with these transitions as depicted for the RON (B), CD44 (C), and ENAH (D) transcripts. H, hnRNP H; M, hnRNP M; MET, mesenchymal–epithelial transition; X, unknown protein.

MYC splicing connections in cancer. As described in the text, MYC acts as master regulator by controlling the expression of splicing factors hnRNP A1, A2, PTBP1, and SRSF1, which promote changes that favor the growth of cancer cells and provide positive feedback loops that further stimulate MYC expression. The KRAS–MAPK pathway, which is relevant to colorectal cancer (KRAS and BRAF mutations) and melanomas (BRAF mutations), converges on the MYC-mediated splicing program through ELK1, which stimulates MYC expression, and Sam68, which improves the production of SRSF1 through alternative splicing. Red arrows are used to indicate impact on alternative splicing.
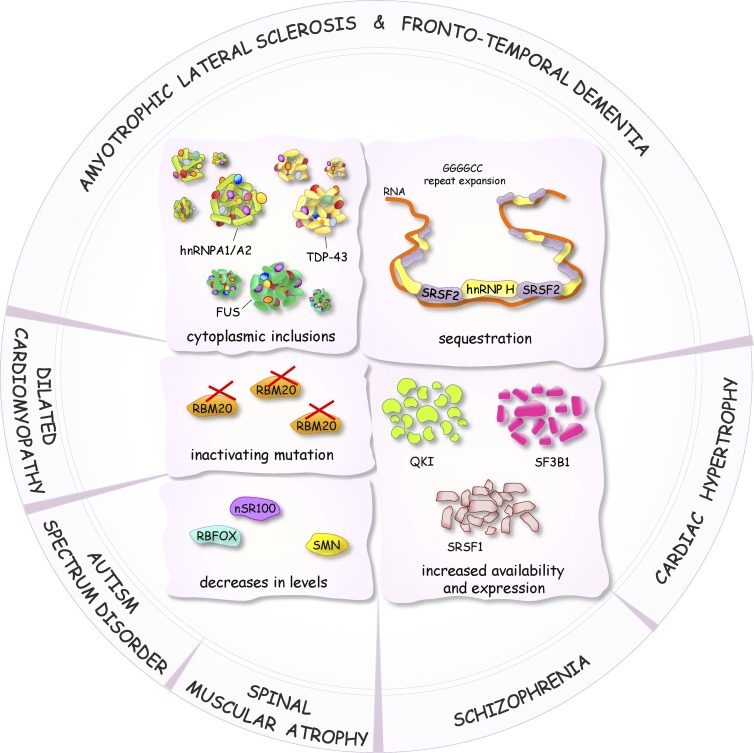
Diagram representing some of the defects in the activity, level, and localization of splicing factors linked to the altered production of splice variants in neurological and muscle-associated diseases.
References
-
- Adler A.S., McCleland M.L., Yee S., Yaylaoglu M., Hussain S., Cosino E., Quinones G., Modrusan Z., Seshagiri S., Torres E., et al. 2014. An integrative analysis of colon cancer identifies an essential function for PRPF6 in tumor growth. Genes Dev. 28:1068–1084. 10.1101/gad.237206.113 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
