

Coevolution of the Organization and Structure of Prokaryotic Genomes
- PMID: 26729648
- PMCID: PMC4691797
- DOI: 10.1101/cshperspect.a018168
Coevolution of the Organization and Structure of Prokaryotic Genomes
Abstract
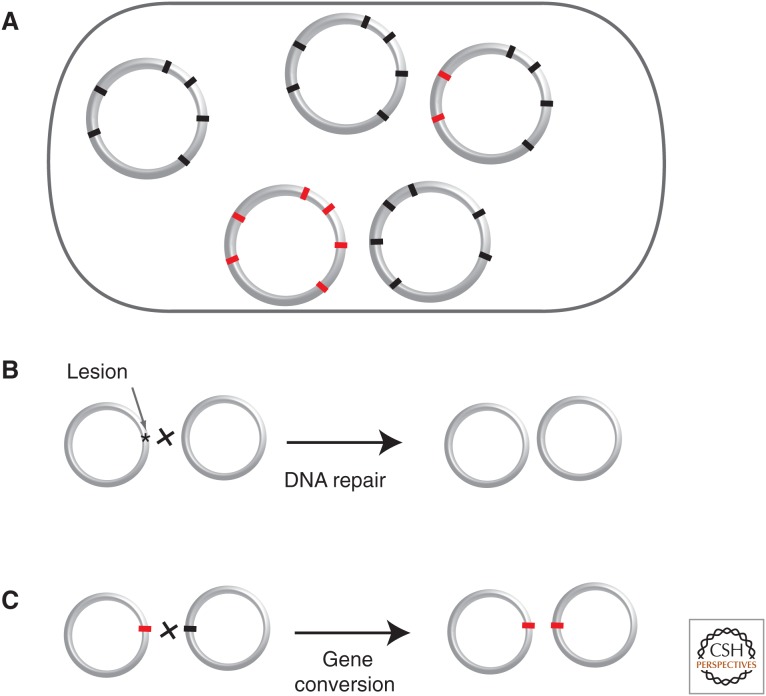
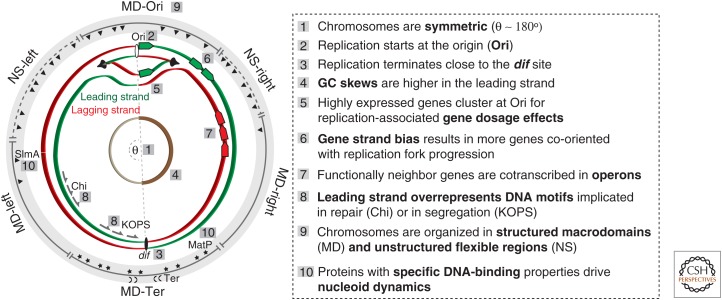
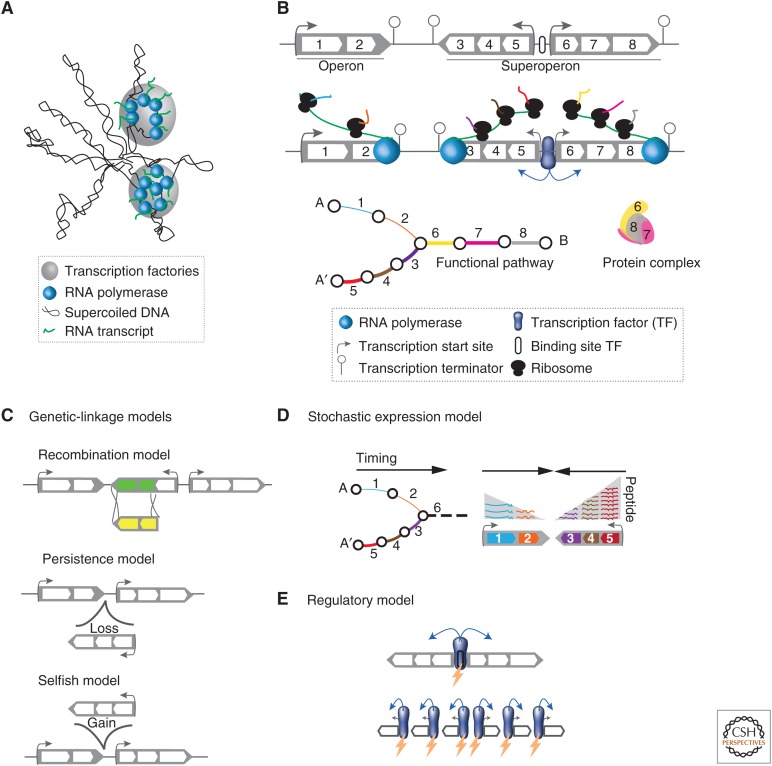
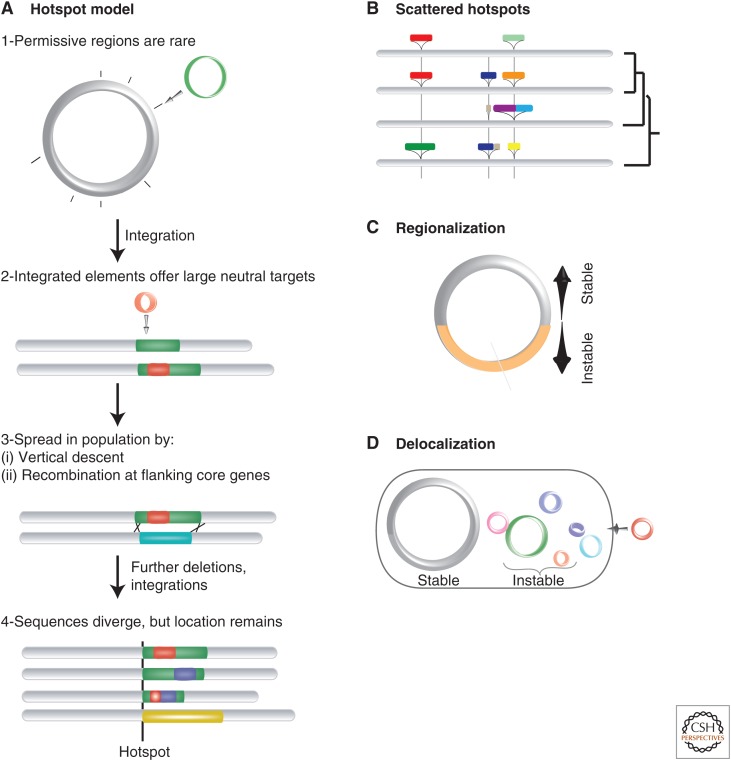
The cytoplasm of prokaryotes contains many molecular machines interacting directly with the chromosome. These vital interactions depend on the chromosome structure, as a molecule, and on the genome organization, as a unit of genetic information. Strong selection for the organization of the genetic elements implicated in these interactions drives replicon ploidy, gene distribution, operon conservation, and the formation of replication-associated traits. The genomes of prokaryotes are also very plastic with high rates of horizontal gene transfer and gene loss. The evolutionary conflicts between plasticity and organization lead to the formation of regions with high genetic diversity whose impact on chromosome structure is poorly understood. Prokaryotic genomes are remarkable documents of natural history because they carry the imprint of all of these selective and mutational forces. Their study allows a better understanding of molecular mechanisms, their impact on microbial evolution, and how they can be tinkered in synthetic biology.
Copyright © 2016 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
-
- Abby S, Daubin V. 2007. Comparative genomics and the evolution of prokaryotes. Trends Microbiol 15: 135–141. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources