

Meiotic DSB patterning: A multifaceted process
- PMID: 26730703
- PMCID: PMC4825777
- DOI: 10.1080/15384101.2015.1093709
Meiotic DSB patterning: A multifaceted process
Abstract
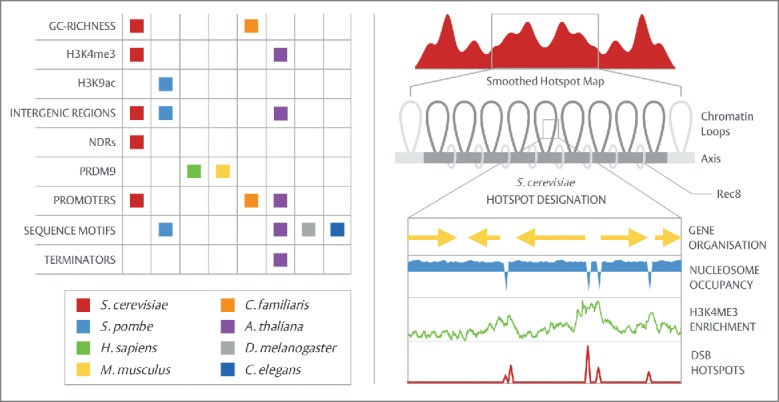
Meiosis is a specialized two-step cell division responsible for genome haploidization and the generation of genetic diversity during gametogenesis. An integral and distinctive feature of the meiotic program is the evolutionarily conserved initiation of homologous recombination (HR) by the developmentally programmed induction of DNA double-strand breaks (DSBs). The inherently dangerous but essential act of DSB formation is subject to multiple forms of stringent and self-corrective regulation that collectively ensure fruitful and appropriate levels of genetic exchange without risk to cellular survival. Within this article we focus upon an emerging element of this control--spatial regulation--detailing recent advances made in understanding how DSBs are evenly distributed across the genome, and present a unified view of the underlying patterning mechanisms employed.
Keywords: ATM; DSB competition; DSB formation; DSB hotspots; DSB interference; Tel1; meiosis; recombination.
Figures



Similar articles
-
Advances Towards How Meiotic Recombination Is Initiated: A Comparative View and Perspectives for Plant Meiosis Research.Int J Mol Sci. 2019 Sep 23;20(19):4718. doi: 10.3390/ijms20194718. Int J Mol Sci. 2019. PMID: 31547623 Free PMC article. Review.
-
Tel1(ATM)-mediated interference suppresses clustered meiotic double-strand-break formation.Nature. 2015 Apr 2;520(7545):114-8. doi: 10.1038/nature13993. Epub 2015 Jan 5. Nature. 2015. PMID: 25539084 Free PMC article.
-
Positive regulation of meiotic DNA double-strand break formation by activation of the DNA damage checkpoint kinase Mec1(ATR).Open Biol. 2013 Jul 31;3(7):130019. doi: 10.1098/rsob.130019. Open Biol. 2013. PMID: 23902647 Free PMC article.
-
Budding yeast ATM/ATR control meiotic double-strand break (DSB) levels by down-regulating Rec114, an essential component of the DSB-machinery.PLoS Genet. 2013 Jun;9(6):e1003545. doi: 10.1371/journal.pgen.1003545. Epub 2013 Jun 27. PLoS Genet. 2013. PMID: 23825959 Free PMC article.
-
The spatial regulation of meiotic recombination hotspots: are all DSB hotspots crossover hotspots?Exp Cell Res. 2012 Jul 15;318(12):1347-52. doi: 10.1016/j.yexcr.2012.03.025. Epub 2012 Mar 31. Exp Cell Res. 2012. PMID: 22487095 Review.
Cited by
-
Chromatin accessibility: methods, mechanisms, and biological insights.Nucleus. 2022 Dec;13(1):236-276. doi: 10.1080/19491034.2022.2143106. Nucleus. 2022. PMID: 36404679 Free PMC article. Review.
-
Regulation Mechanisms of Meiotic Recombination Revealed from the Analysis of a Fission Yeast Recombination Hotspot ade6-M26.Biomolecules. 2022 Nov 26;12(12):1761. doi: 10.3390/biom12121761. Biomolecules. 2022. PMID: 36551189 Free PMC article. Review.
-
The roles of homologous recombination and the immune system in the genomic evolution of cancer.J Transl Sci. 2019 Apr;5(2):10.15761/JTS.1000282. doi: 10.15761/JTS.1000282. Epub 2018 Oct 1. J Transl Sci. 2019. PMID: 30873294 Free PMC article.
-
DNA-driven condensation assembles the meiotic DNA break machinery.Nature. 2021 Apr;592(7852):144-149. doi: 10.1038/s41586-021-03374-w. Epub 2021 Mar 17. Nature. 2021. PMID: 33731927 Free PMC article.
-
The CNOT4 Subunit of the CCR4-NOT Complex is Involved in mRNA Degradation, Efficient DNA Damage Repair, and XY Chromosome Crossover during Male Germ Cell Meiosis.Adv Sci (Weinh). 2021 Mar 16;8(10):2003636. doi: 10.1002/advs.202003636. eCollection 2021 May. Adv Sci (Weinh). 2021. PMID: 34026442 Free PMC article.
References
-
- Lam I, Keeney S. Mechanism and regulation of meiotic recombination initiation. Cold Spring Harb Perspect Biol 2015; 7:a016634; http://dx.doi.org/10.1101/cshperspect.a016634 - DOI - PMC - PubMed
-
- Keeney S, Lange J, Mohibullah N. Self-organization of meiotic recombination initiation: general principles and molecular pathways. Annu Rev Genet 2014; 48:187-214; PMID:25421598; http://dx.doi.org/10.1146/annurev-genet-120213-092304 - DOI - PMC - PubMed
-
- Khil PP, Smagulova F, Brick KM, Camerini-Otero RD, Petukhova GV. Sensitive mapping of recombination hotspots using sequencing-based detection of ssDNA. Genome Res 2012; 22:957-65; PMID:22367190; http://dx.doi.org/10.1101/gr.130583.111 - DOI - PMC - PubMed
-
- Pan J, Sasaki M, Kniewel R, Murakami H, Blitzblau HG, Tischfield SE, Zhu X, Neale MJ, Jasin M, Socci ND, et al. . A hierarchical combination of factors shapes the genome-wide topography of yeast meiotic recombination initiation. Cell 2011; 144:719-31; PMID:21376234; http://dx.doi.org/10.1016/j.cell.2011.02.009 - DOI - PMC - PubMed
-
- Pratto F, Brick K, Khil P, Smagulova F, Petukhova GV, Camerini-Otero RD. Recombination initiation maps of individual human genomes. Science 2014; 346:1256442-2; PMID:25395542; http://dx.doi.org/10.1126/science.1256442 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous