

The TIM Barrel Architecture Facilitated the Early Evolution of Protein-Mediated Metabolism
- PMID: 26733481
- PMCID: PMC4709378
- DOI: 10.1007/s00239-015-9722-8
The TIM Barrel Architecture Facilitated the Early Evolution of Protein-Mediated Metabolism
Abstract
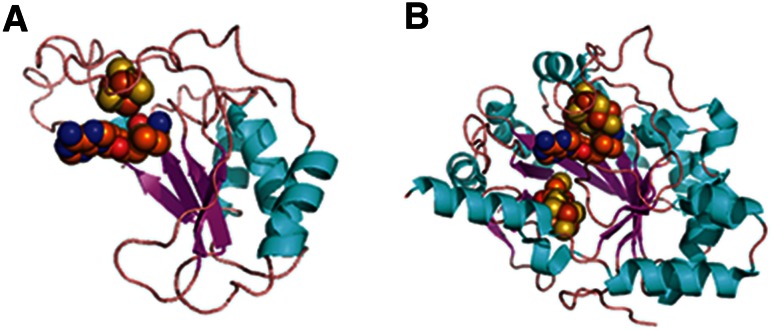
The triosephosphate isomerase (TIM) barrel protein fold is a structurally repetitive architecture that is present in approximately 10% of all enzymes. It is generally assumed that this ubiquity in modern proteomes reflects an essential historical role in early protein-mediated metabolism. Here, we provide quantitative and comparative analyses to support several hypotheses about the early importance of the TIM barrel architecture. An information theoretical analysis of protein structures supports the hypothesis that the TIM barrel architecture could arise more easily by duplication and recombination compared to other mixed α/β structures. We show that TIM barrel enzymes corresponding to the most taxonomically broad superfamilies also have the broadest range of functions, often aided by metal and nucleotide-derived cofactors that are thought to reflect an earlier stage of metabolic evolution. By comparison to other putatively ancient protein architectures, we find that the functional diversity of TIM barrel proteins cannot be explained simply by their antiquity. Instead, the breadth of TIM barrel functions can be explained, in part, by the incorporation of a broad range of cofactors, a trend that does not appear to be shared by proteins in general. These results support the hypothesis that the simple and functionally general TIM barrel architecture may have arisen early in the evolution of protein biosynthesis and provided an ideal scaffold to facilitate the metabolic transition from ribozymes, peptides, and geochemical catalysts to modern protein enzymes.
Keywords: LUCA; Prebiotic chemistry; RNA world; RNA–protein world; TIM barrel.
Figures
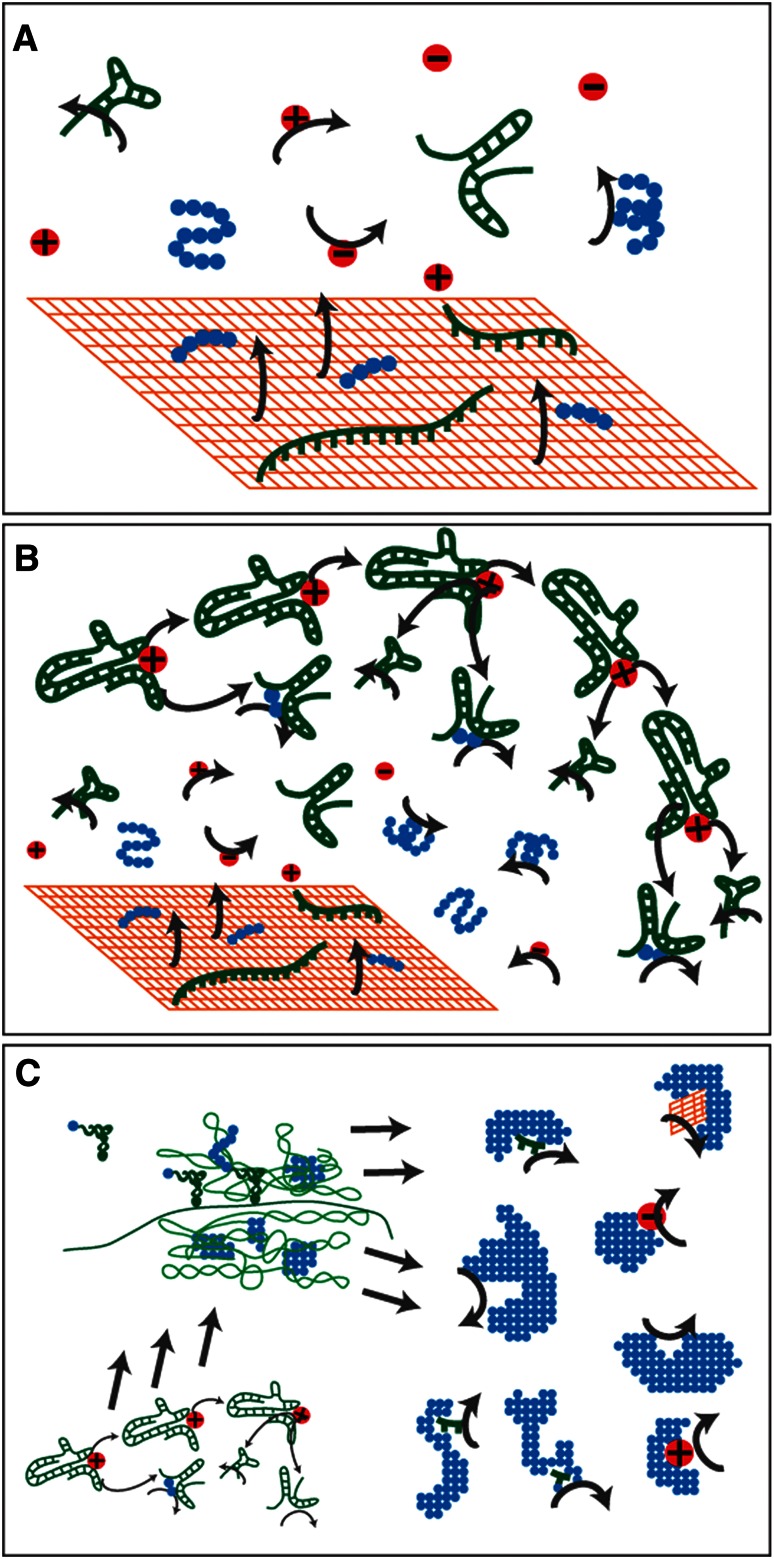
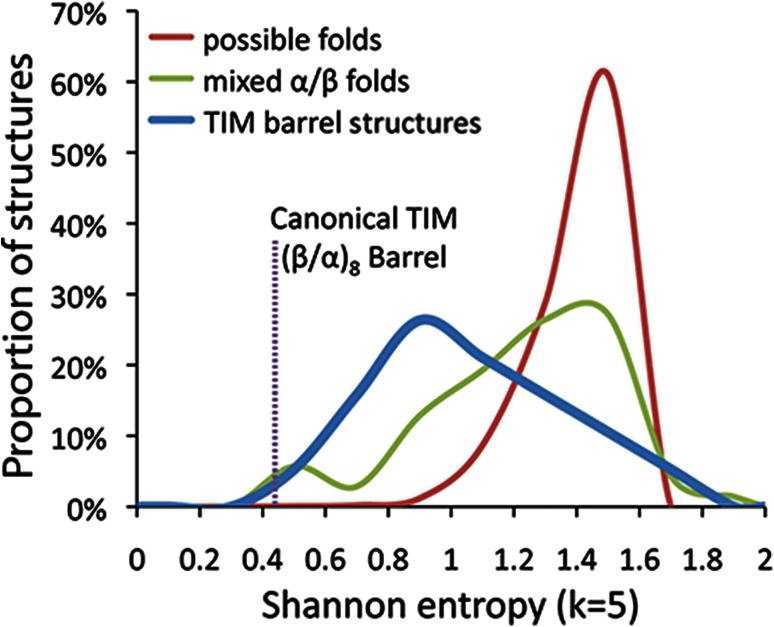
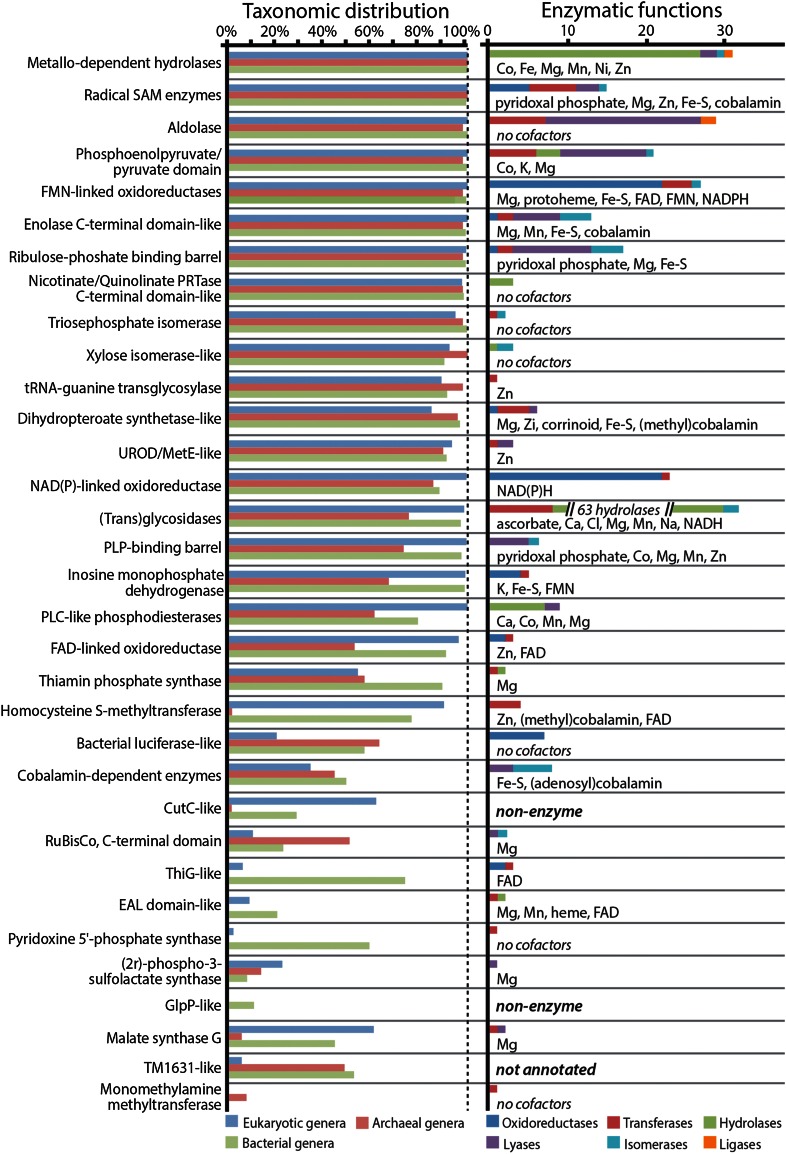
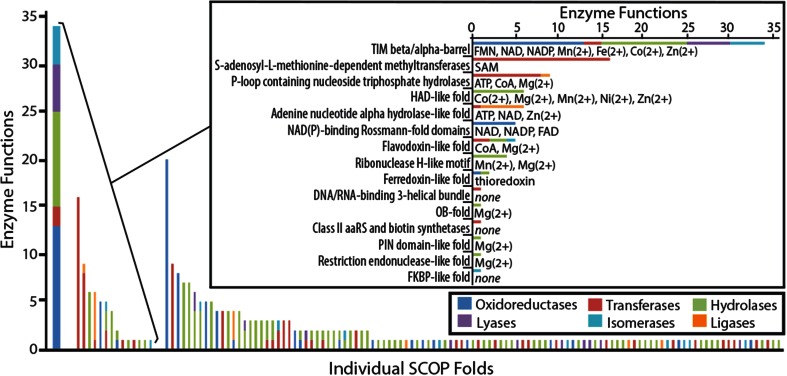
References
-
- Becerra A, Delaye L, Islas S, Lazcano A. The very early stages of biological evolution and the nature of the last common ancestor of the three major cell domains. Annu Rev Ecol Evol Syst. 2007;38:361–379. doi: 10.1146/annurev.ecolsys.38.091206.095825. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
