

Extracardiac septum transversum/proepicardial endothelial cells pattern embryonic coronary arterio-venous connections
- PMID: 26739565
- PMCID: PMC4725486
- DOI: 10.1073/pnas.1509834113
Extracardiac septum transversum/proepicardial endothelial cells pattern embryonic coronary arterio-venous connections
Abstract
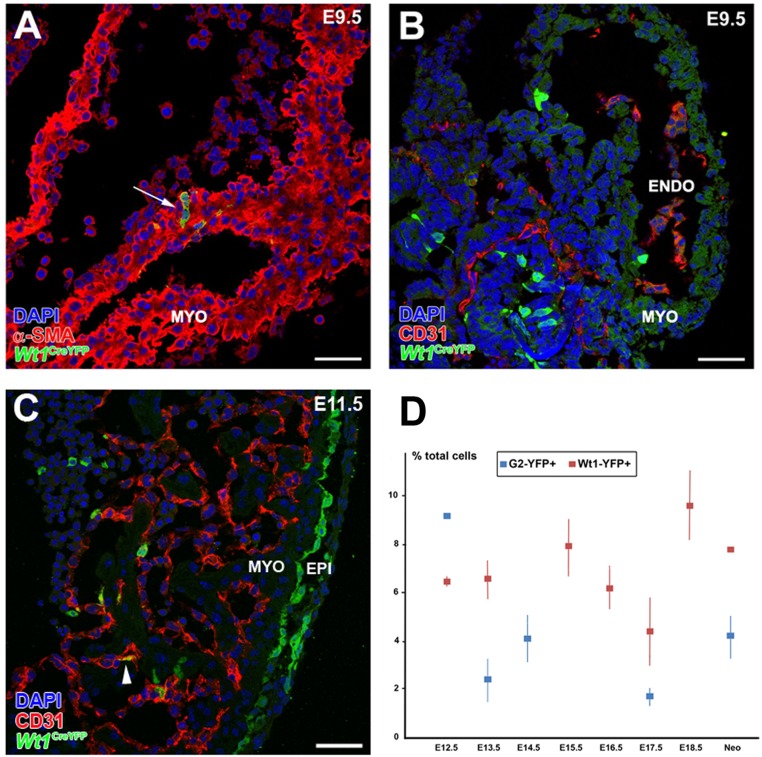
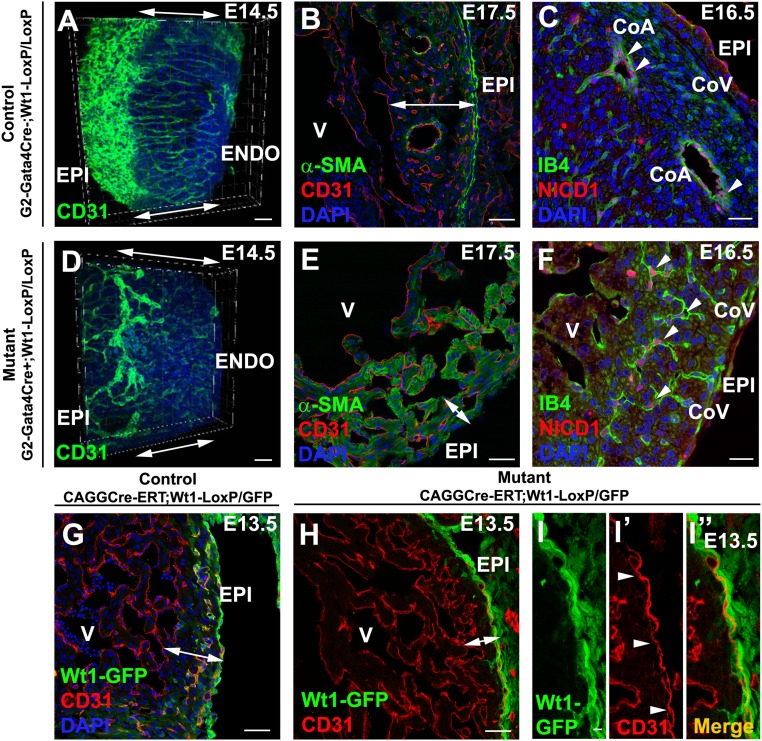
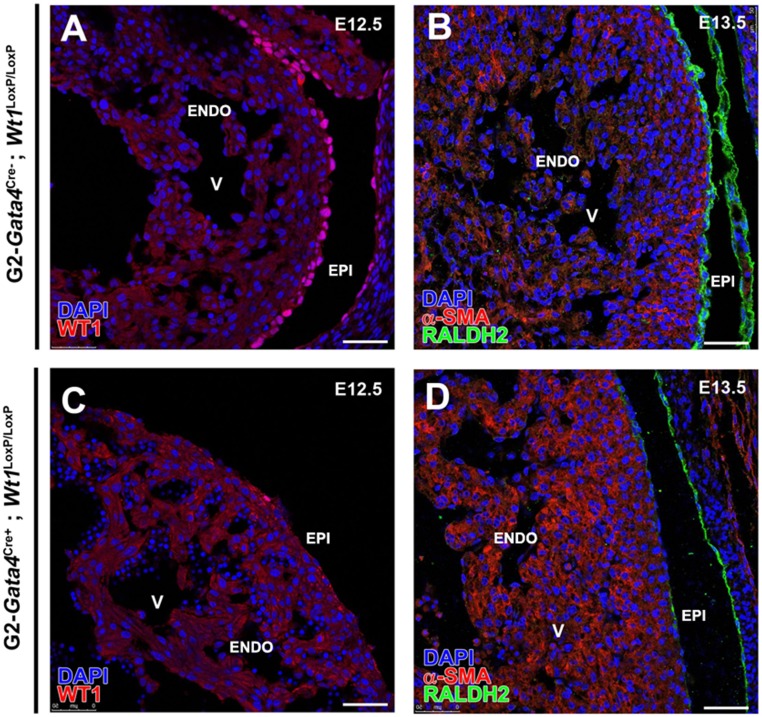
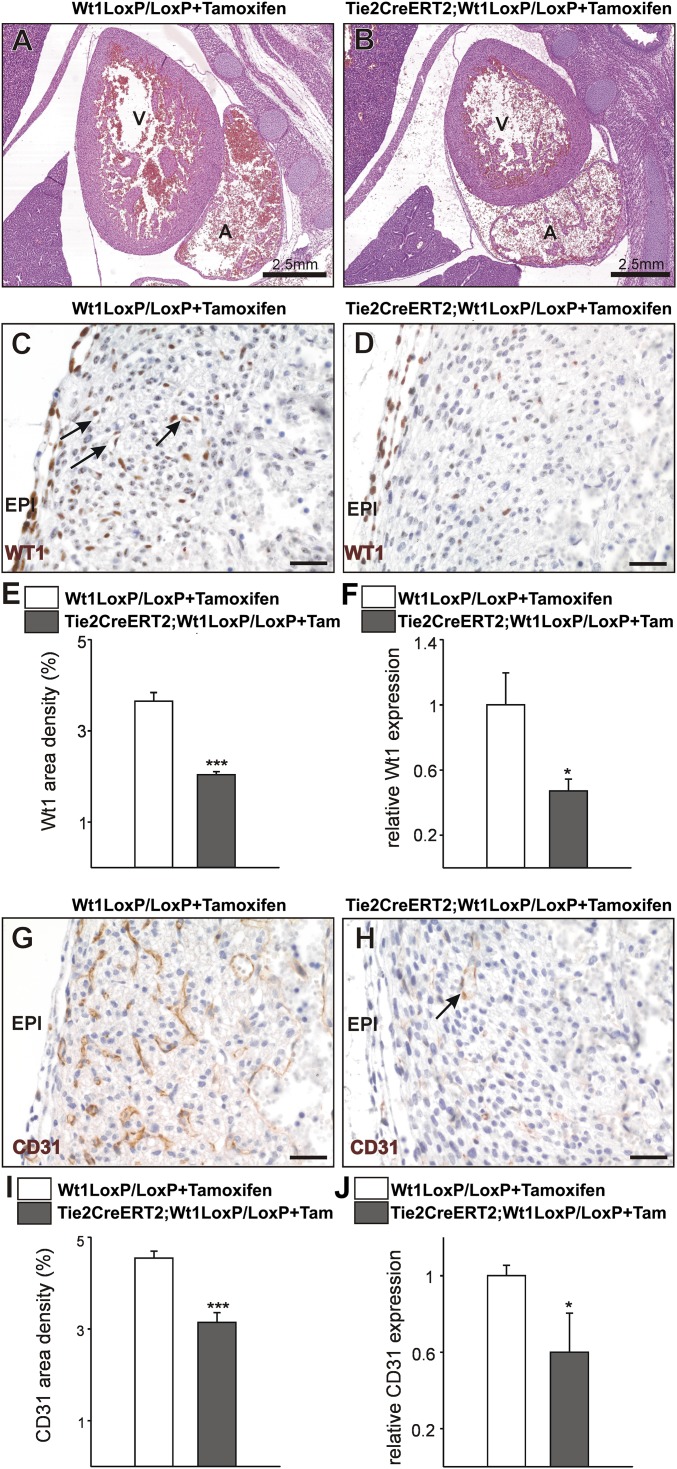
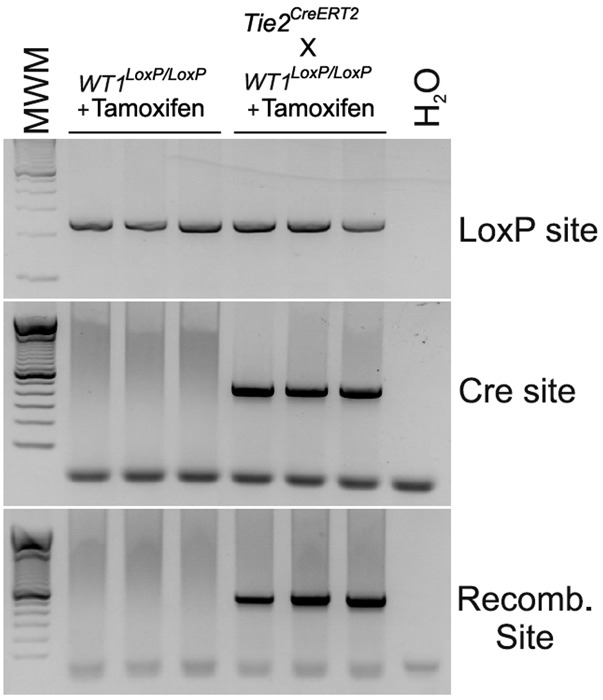
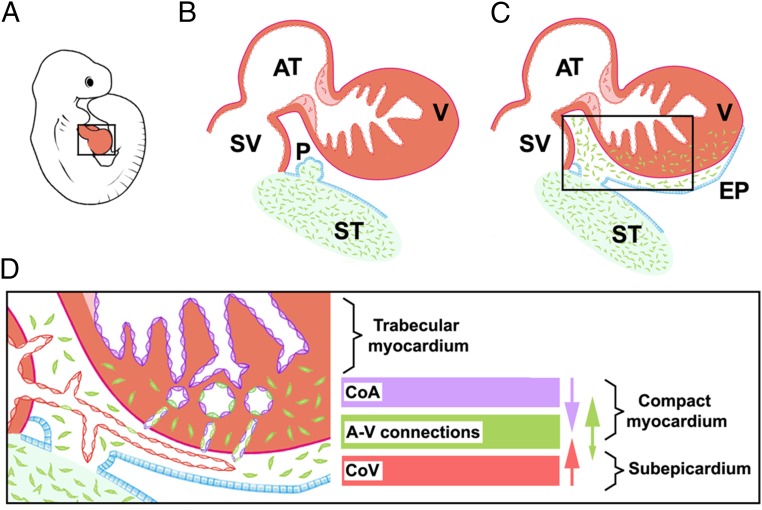
Recent reports suggest that mammalian embryonic coronary endothelium (CoE) originates from the sinus venosus and ventricular endocardium. However, the contribution of extracardiac cells to CoE is thought to be minor and nonsignificant for coronary formation. Using classic (Wt1(Cre)) and previously undescribed (G2-Gata4(Cre)) transgenic mouse models for the study of coronary vascular development, we show that extracardiac septum transversum/proepicardium (ST/PE)-derived endothelial cells are required for the formation of ventricular coronary arterio-venous vascular connections. Our results indicate that at least 20% of embryonic coronary arterial and capillary endothelial cells derive from the ST/PE compartment. Moreover, we show that conditional deletion of the ST/PE lineage-specific Wilms' tumor suppressor gene (Wt1) in the ST/PE of G2-Gata4(Cre) mice and in the endothelium of Tie2(Cre) mice disrupts embryonic coronary transmural patterning, leading to embryonic death. Taken together, our results demonstrate that ST/PE-derived endothelial cells contribute significantly to and are required for proper coronary vascular morphogenesis.
Keywords: Gata4; Wt1; coronary endothelium; proepicardium; septum transversum.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
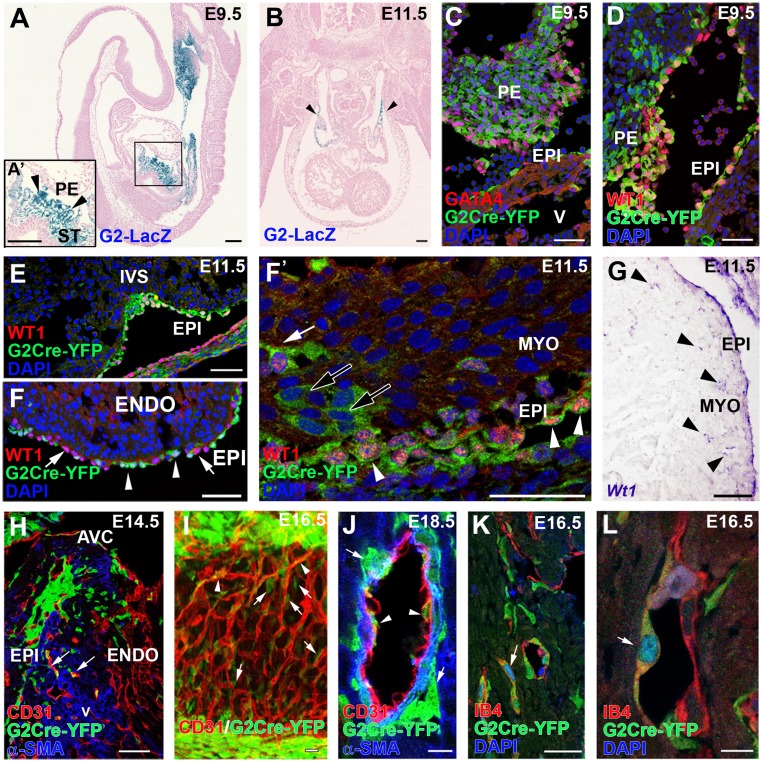
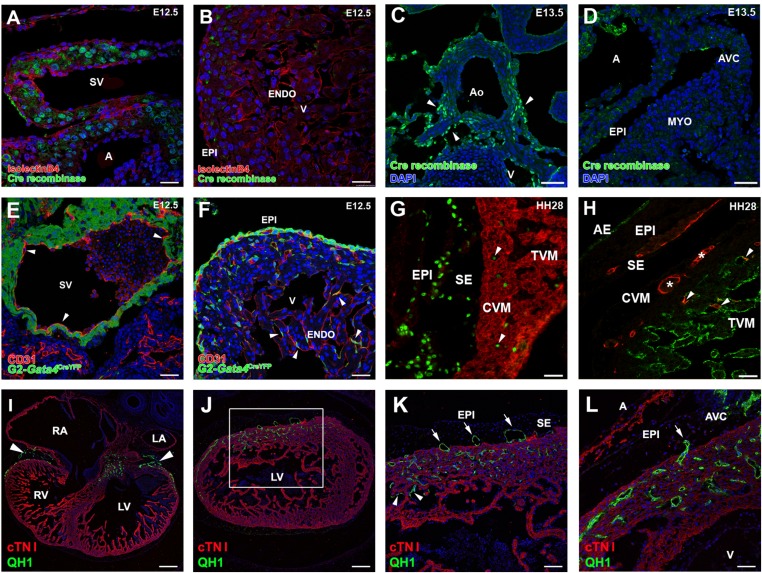
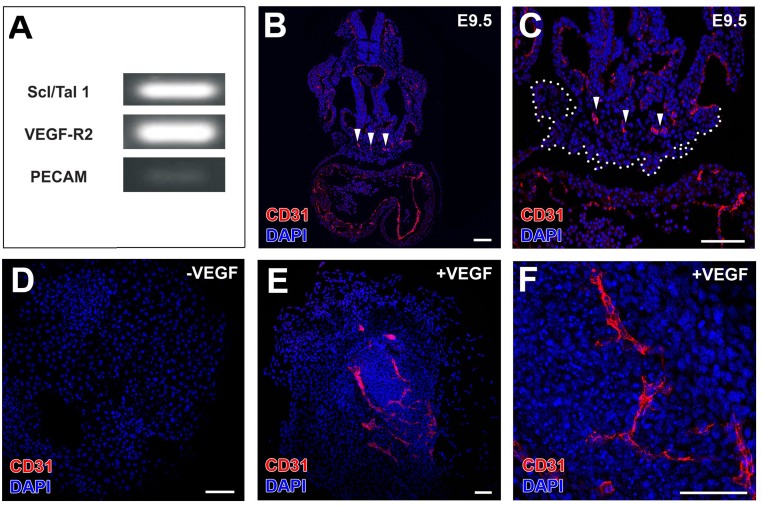
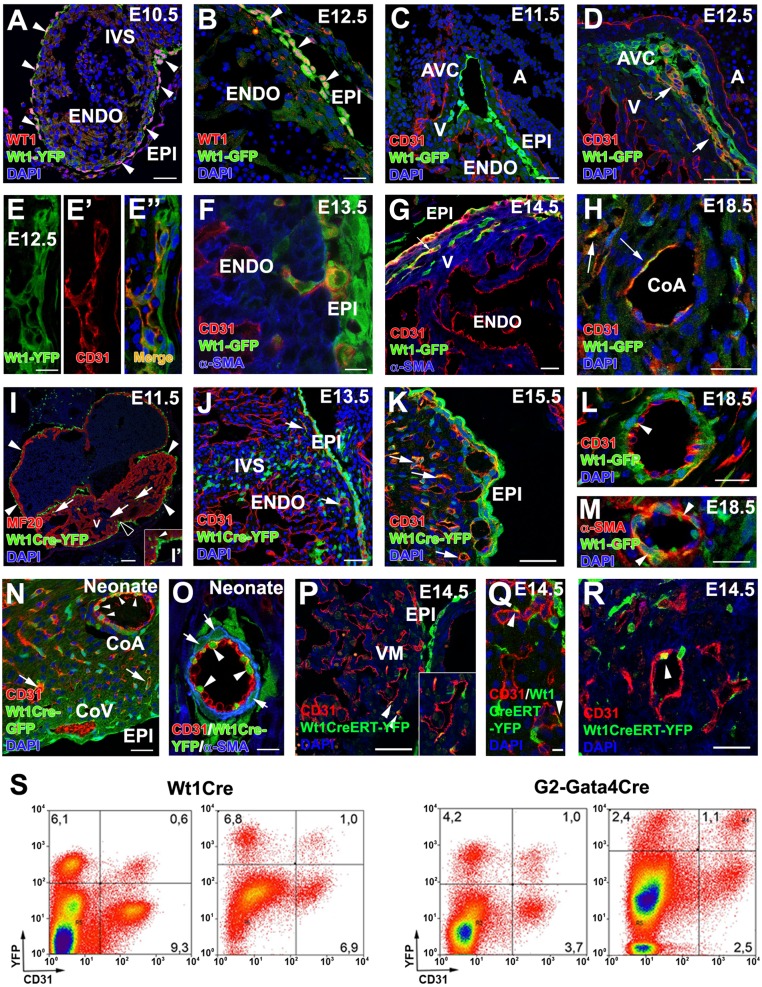
References
-
- Pérez-Pomares JM, de la Pompa JL. Signaling during epicardium and coronary vessel development. Circ Res. 2011;109(12):1429–1442. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
