

Hippocampal Respiration-Driven Rhythm Distinct from Theta Oscillations in Awake Mice
- PMID: 26740658
- PMCID: PMC6601786
- DOI: 10.1523/JNEUROSCI.2848-15.2016
Hippocampal Respiration-Driven Rhythm Distinct from Theta Oscillations in Awake Mice
Abstract
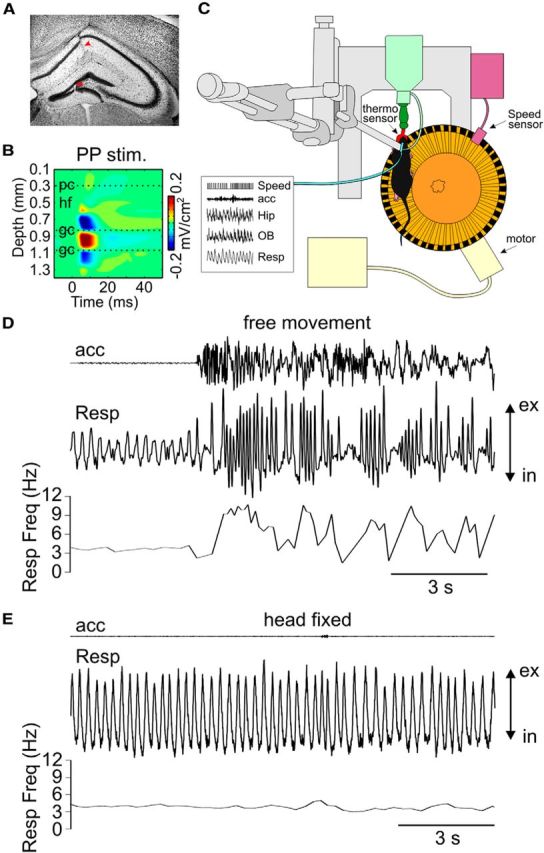
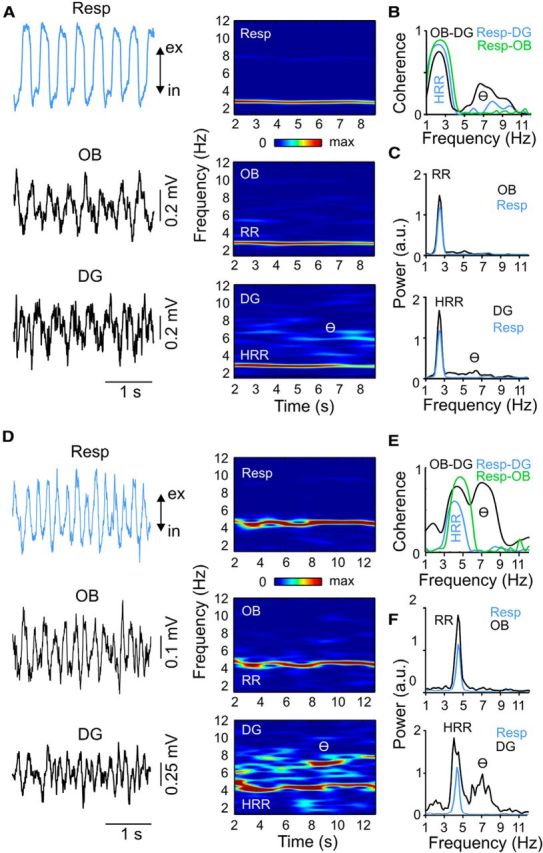
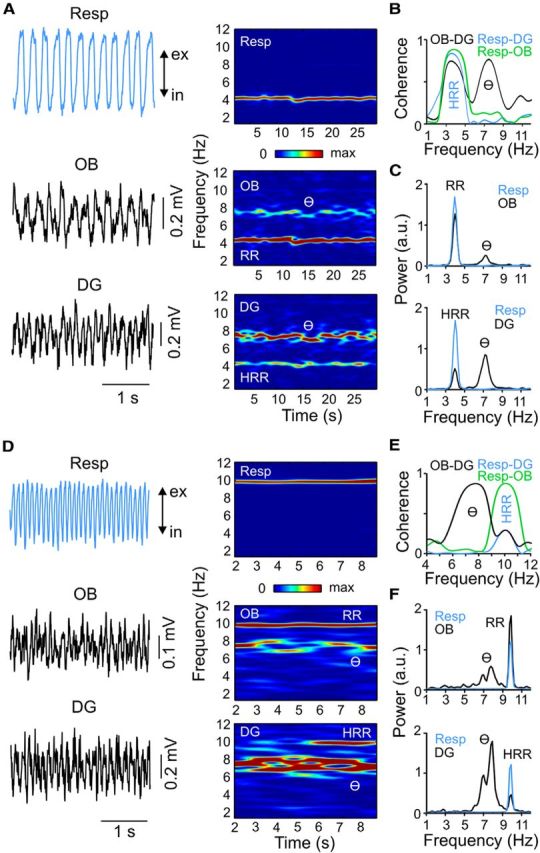
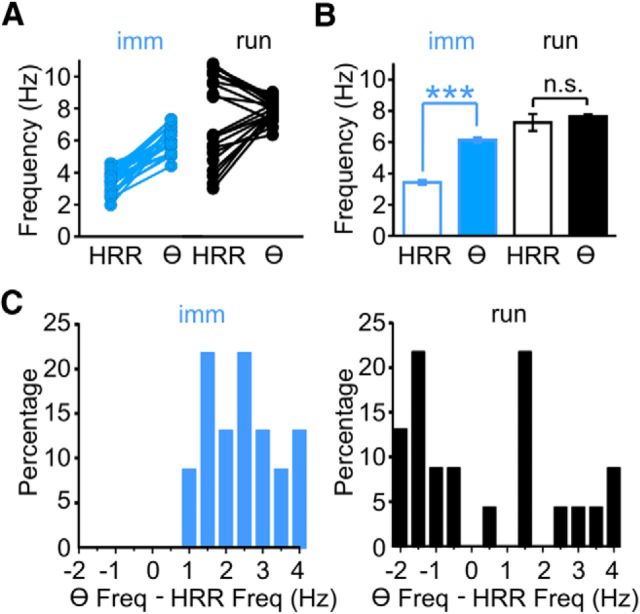
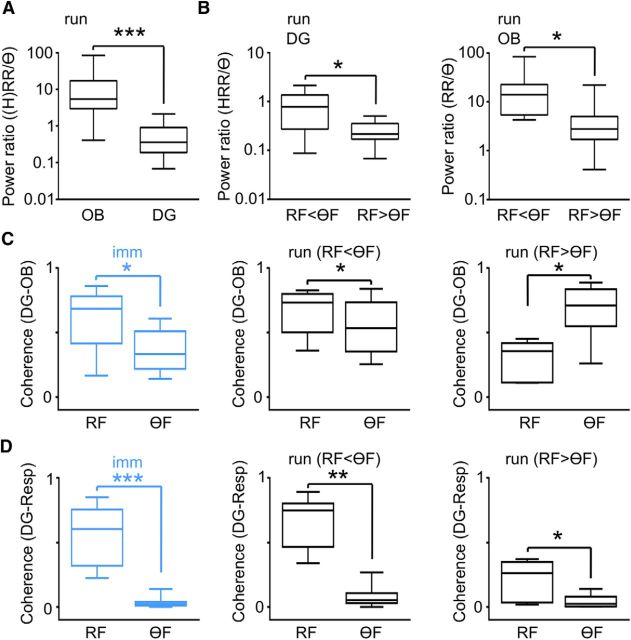
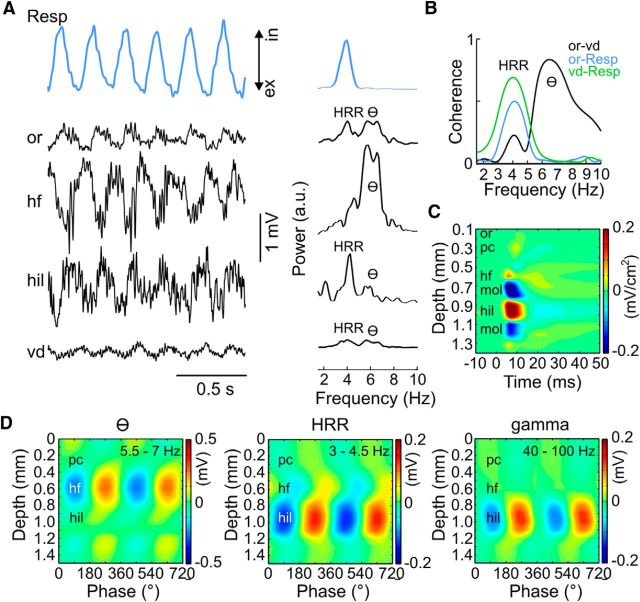
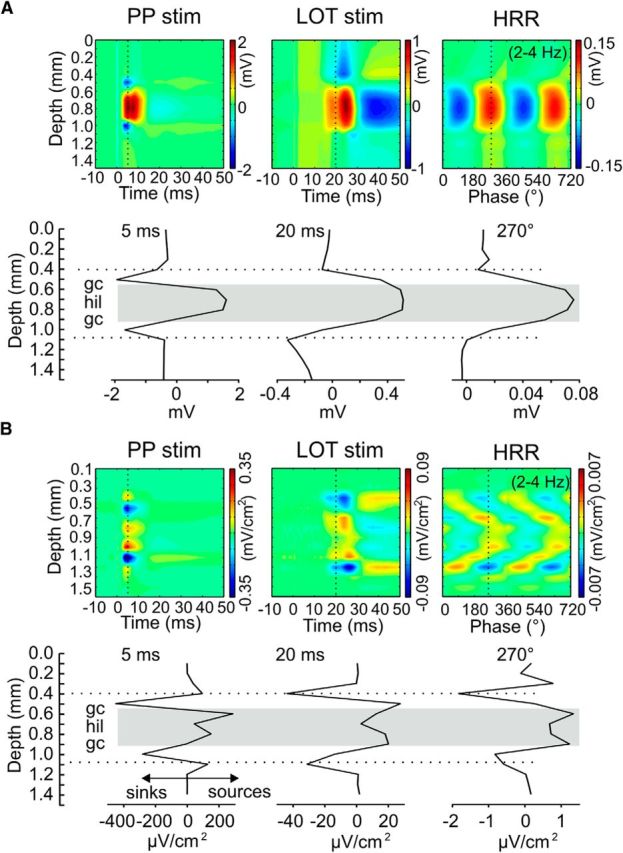
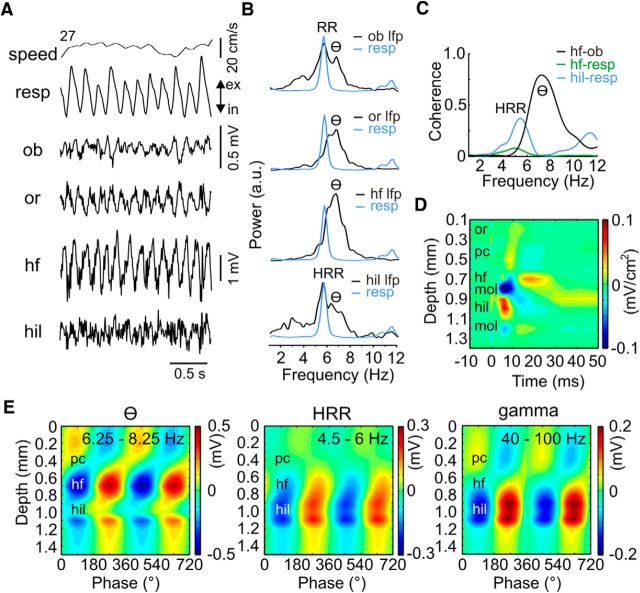
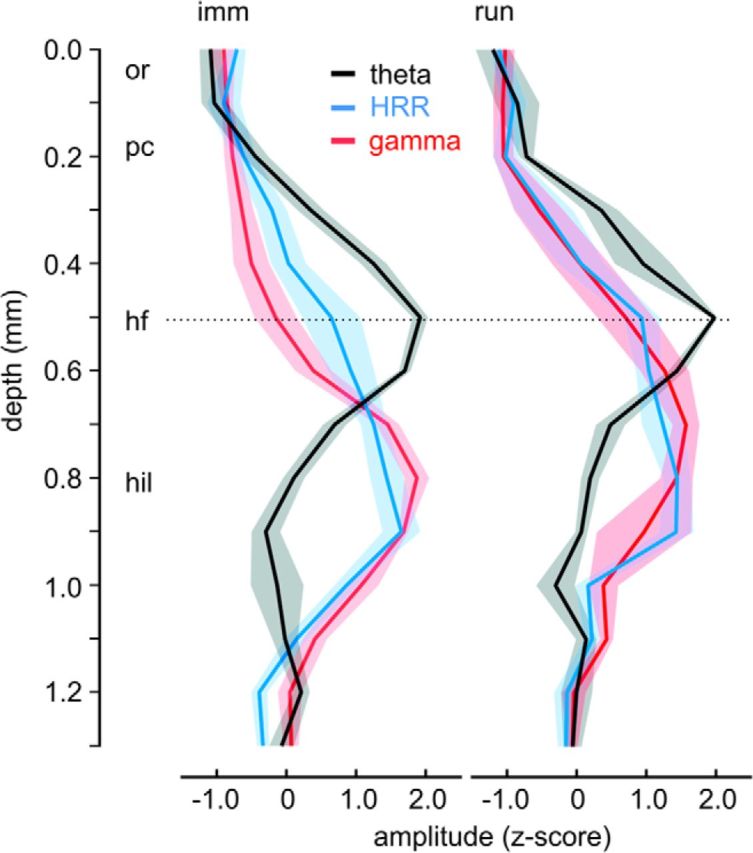
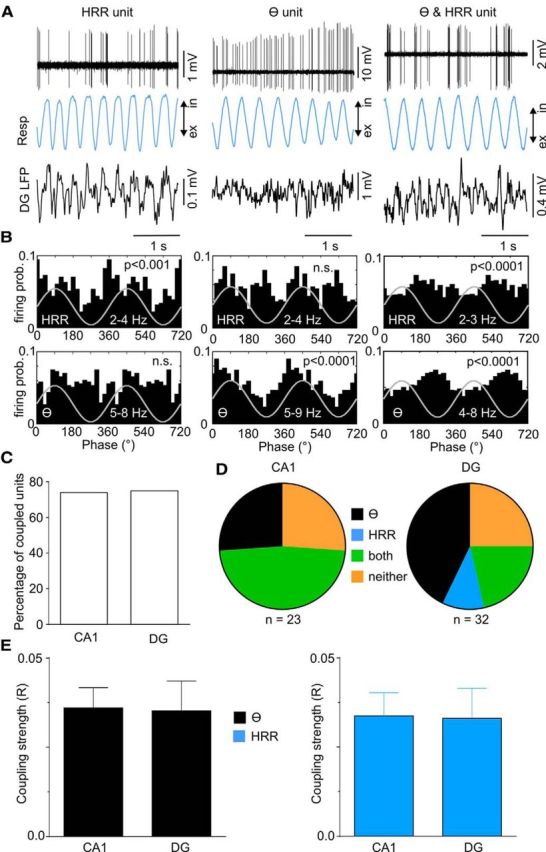
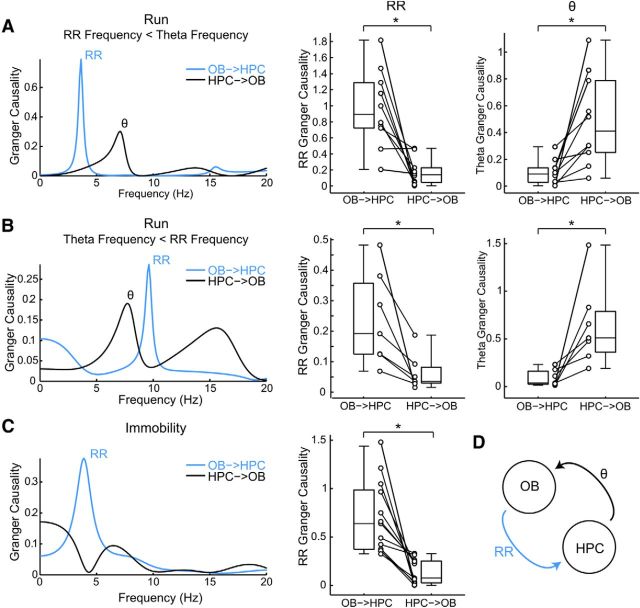
We have recently described a slow oscillation in the hippocampus of urethane-anesthetized mice, which couples to nasal respiration and is clearly distinct from co-occurring theta oscillations. Here we set out to investigate whether such type of patterned network activity, which we named "hippocampal respiration rhythm" (HRR), also occurs in awake mice. In freely moving mice, instantaneous respiration rate is extremely variable, and respiration is superimposed by bouts of sniffing. To reduce this variability, we clamped the behavior of the animal to either awake immobility or treadmill running by using a head-fixed setup while simultaneously recording respiration and field potentials from the olfactory bulb (OB) and hippocampus. Head-fixed animals often exhibited long periods of steady respiration rate during either immobility or running, which allowed for spectral and coherence analyses with a sufficient frequency resolution to sort apart respiration and theta activities. We could thus demonstrate the existence of HRR in awake animals, namely, a respiration-entrained slow rhythm with highest amplitude at the dentate gyrus. HRR was most prominent during immobility and running with respiration rates slower than theta oscillations. Nevertheless, HRR could also be faster than theta. Discharges of juxtacellularly recorded cells in CA1 and dentate gyrus were modulated by HRR and theta oscillations. Granger directionality analysis revealed that HRR is caused by the OB and that theta oscillations in OB are caused by the hippocampus. Our results suggest that respiration-coupled oscillations aid the exchange of information between olfactory and memory networks.
Significance statement: Olfaction is a major sense in rodents. In consequence, the olfactory bulb (OB) should be able to transmit information to downstream regions. Here we report potential mechanisms underlying such information transfer. We demonstrate the existence of a respiration-entrained rhythm in the hippocampus of awake mice. Frequencies of the hippocampal respiration rhythm (HRR) overlap with classical theta oscillations, but both rhythms are clearly distinct. HRR is most prominent in the dentate gyrus, especially when respiration is slower than theta frequency. Discharges of neurons in CA1 and dentate gyrus are modulated by both HRR and theta. Directionality analysis shows that HRR is caused by the OB. Our results suggest that respiration-coupled oscillations aid the exchange of information between olfactory and memory networks.
Keywords: hippocampus; mouse; olfactory bulb; oscillation; respiration; theta rhythm.
Copyright © 2016 the authors 0270-6474/16/360162-16$15.00/0.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous