

Sustained Suppression of Hyperalgesia during Latent Sensitization by μ-, δ-, and κ-opioid receptors and α2A Adrenergic Receptors: Role of Constitutive Activity
- PMID: 26740662
- PMCID: PMC4701961
- DOI: 10.1523/JNEUROSCI.1751-15.2016
Sustained Suppression of Hyperalgesia during Latent Sensitization by μ-, δ-, and κ-opioid receptors and α2A Adrenergic Receptors: Role of Constitutive Activity
Abstract
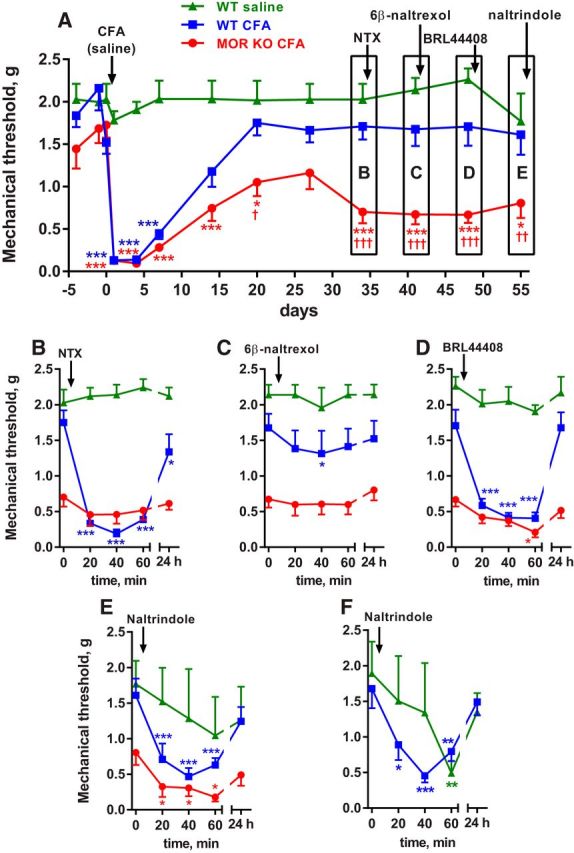
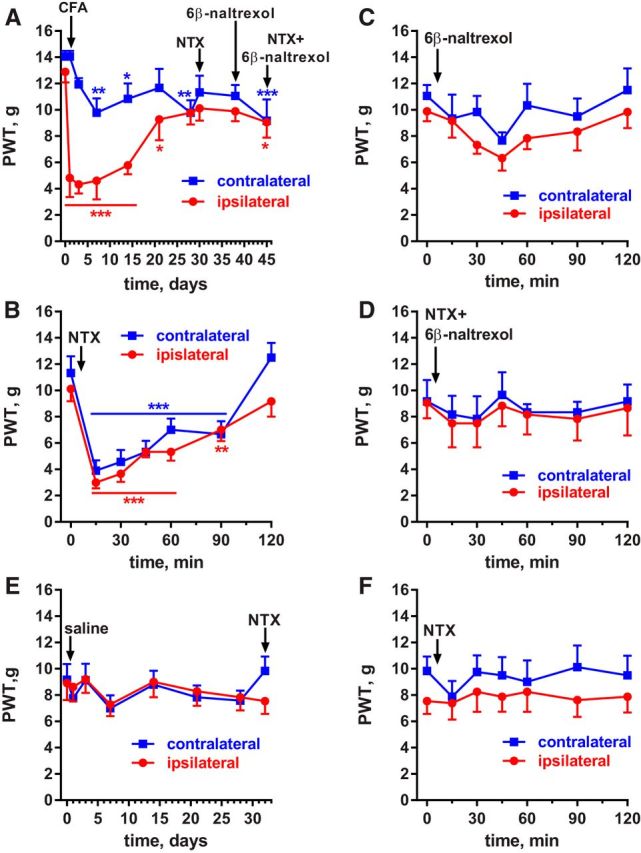
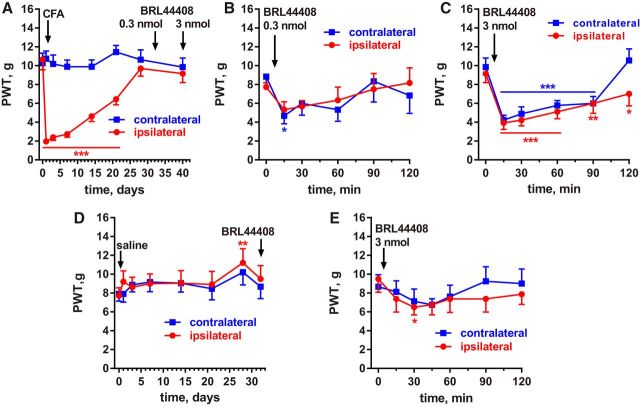
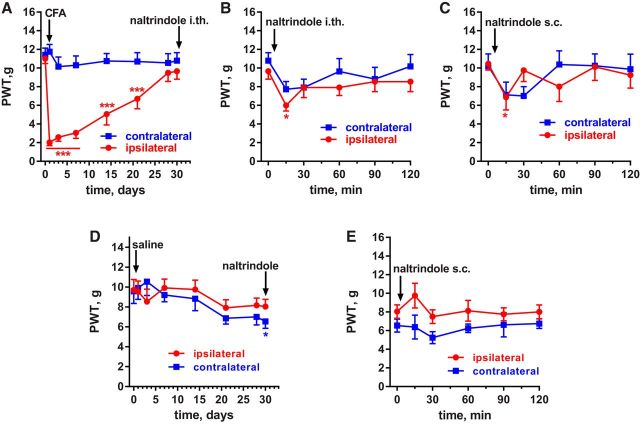
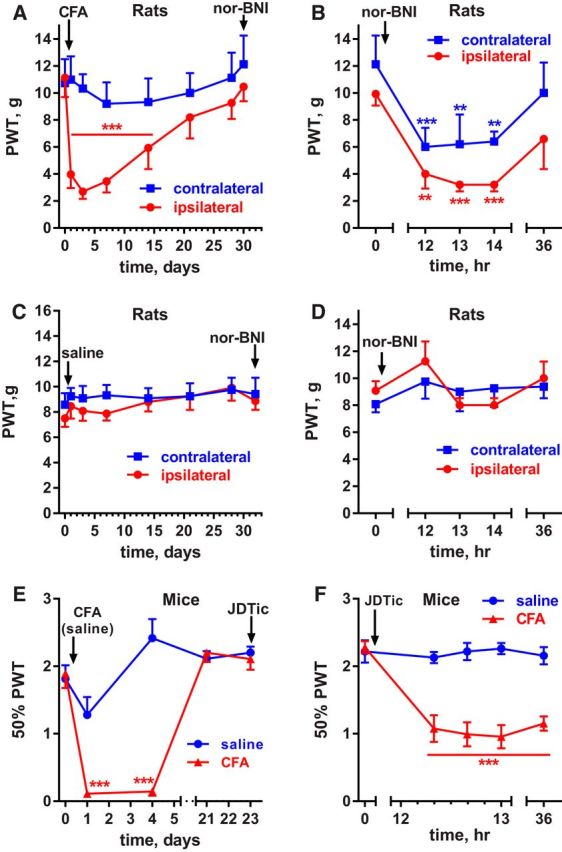
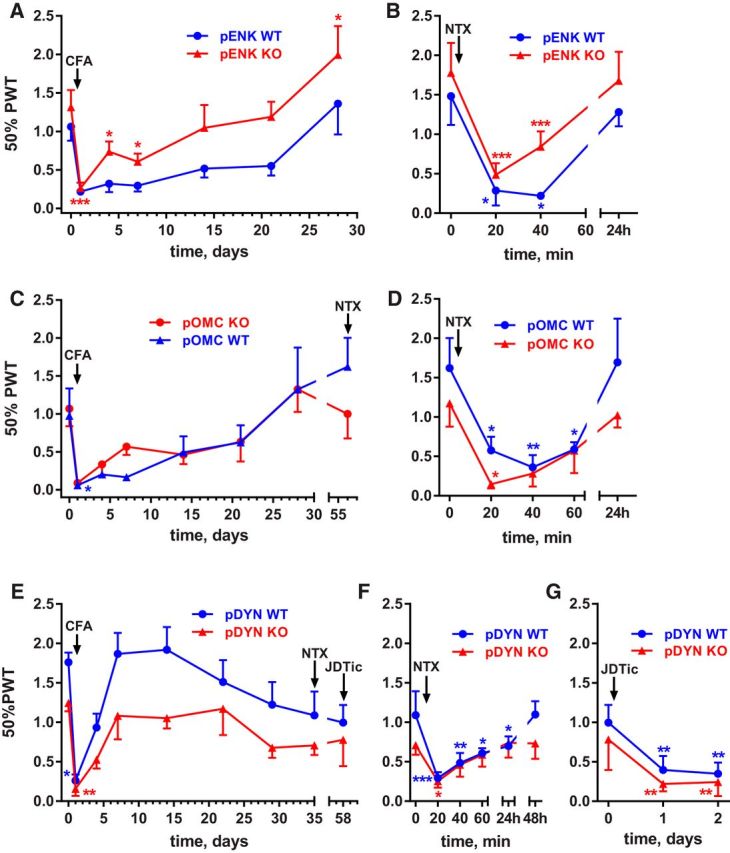
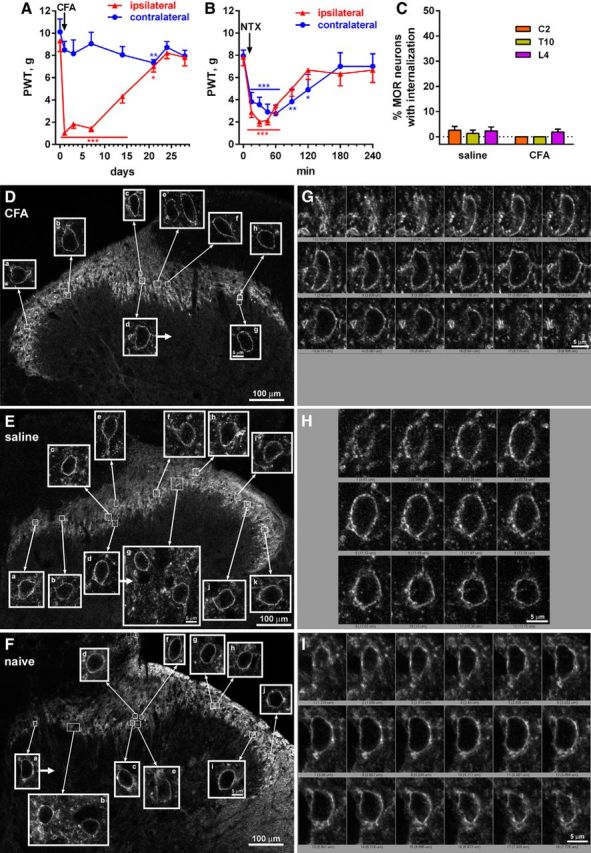
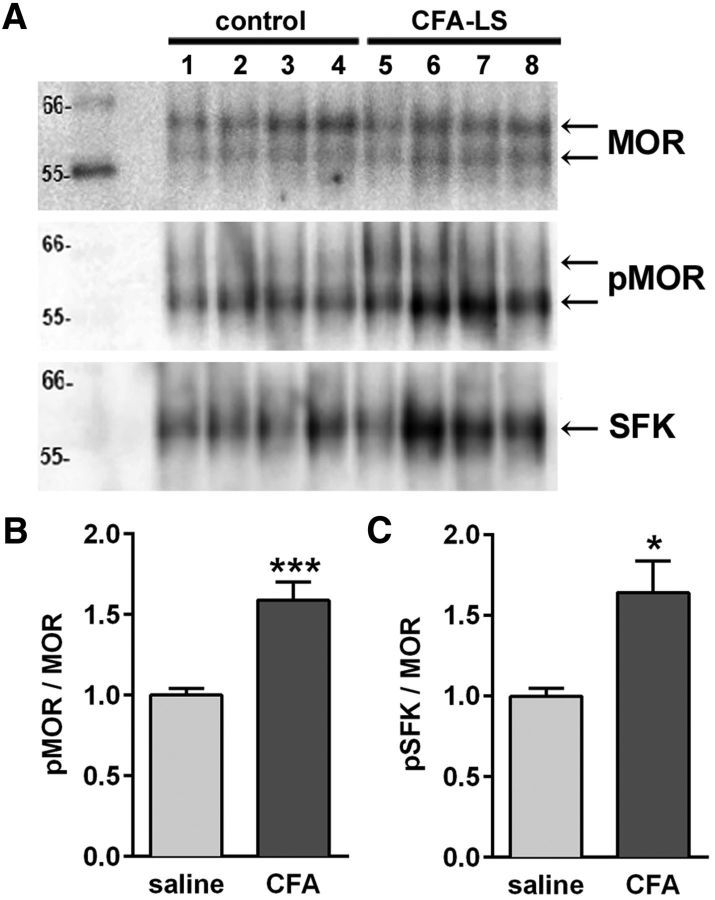
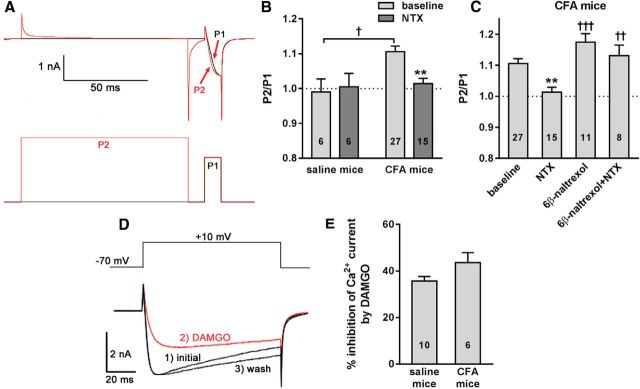
Many chronic pain disorders alternate between bouts of pain and periods of remission. The latent sensitization model reproduces this in rodents by showing that the apparent recovery ("remission") from inflammatory or neuropathic pain can be reversed by opioid antagonists. Therefore, this remission represents an opioid receptor-mediated suppression of a sustained hyperalgesic state. To identify the receptors involved, we induced latent sensitization in mice and rats by injecting complete Freund's adjuvant (CFA) in the hindpaw. In WT mice, responses to mechanical stimulation returned to baseline 3 weeks after CFA. In μ-opioid receptor (MOR) knock-out (KO) mice, responses did not return to baseline but partially recovered from peak hyperalgesia. Antagonists of α2A-adrenergic and δ-opioid receptors reinstated hyperalgesia in WT mice and abolished the partial recovery from hyperalgesia in MOR KO mice. In rats, antagonists of α2A adrenergic and μ-, δ-, and κ-opioid receptors reinstated hyperalgesia during remission from CFA-induced hyperalgesia. Therefore, these four receptors suppress hyperalgesia in latent sensitization. We further demonstrated that suppression of hyperalgesia by MORs was due to their constitutive activity because of the following: (1) CFA-induced hyperalgesia was reinstated by the MOR inverse agonist naltrexone (NTX), but not by its neutral antagonist 6β-naltrexol; (2) pro-enkephalin, pro-opiomelanocortin, and pro-dynorphin KO mice showed recovery from hyperalgesia and reinstatement by NTX; (3) there was no MOR internalization during remission; (4) MORs immunoprecipitated from the spinal cord during remission had increased Ser(375) phosphorylation; and (5) electrophysiology recordings from dorsal root ganglion neurons collected during remission showed constitutive MOR inhibition of calcium channels.
Significance statement: Chronic pain causes extreme suffering to millions of people, but its mechanisms remain to be unraveled. Latent sensitization is a phenomenon studied in rodents that has many key features of chronic pain: it is initiated by a variety of noxious stimuli, has indefinite duration, and pain appears in episodes that can be triggered by stress. Here, we show that, during latent sensitization, there is a sustained state of pain hypersensitivity that is continuously suppressed by the activation of μ-, δ-, and κ-opioid receptors and by adrenergic α2A receptors in the spinal cord. Furthermore, we show that the activation of μ-opioid receptors is not due to the release of endogenous opioids, but rather to its ligand-independent constitutive activity.
Keywords: adrenergic receptor; constitutive activity; hyperalgesia; kappa-opioid receptor; latent sensitization; mu-opioid receptor.
Copyright © 2016 the authors 0270-6474/16/360204-18$15.00/0.
Figures
Comment in
-
PC-FACS.J Pain Symptom Manage. 2016 Jun;51(6):1105-11. doi: 10.1016/j.jpainsymman.2016.05.001. Epub 2016 May 12. J Pain Symptom Manage. 2016. PMID: 27179560
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials