

TIGIT Marks Exhausted T Cells, Correlates with Disease Progression, and Serves as a Target for Immune Restoration in HIV and SIV Infection
- PMID: 26741490
- PMCID: PMC4704737
- DOI: 10.1371/journal.ppat.1005349
TIGIT Marks Exhausted T Cells, Correlates with Disease Progression, and Serves as a Target for Immune Restoration in HIV and SIV Infection
Abstract
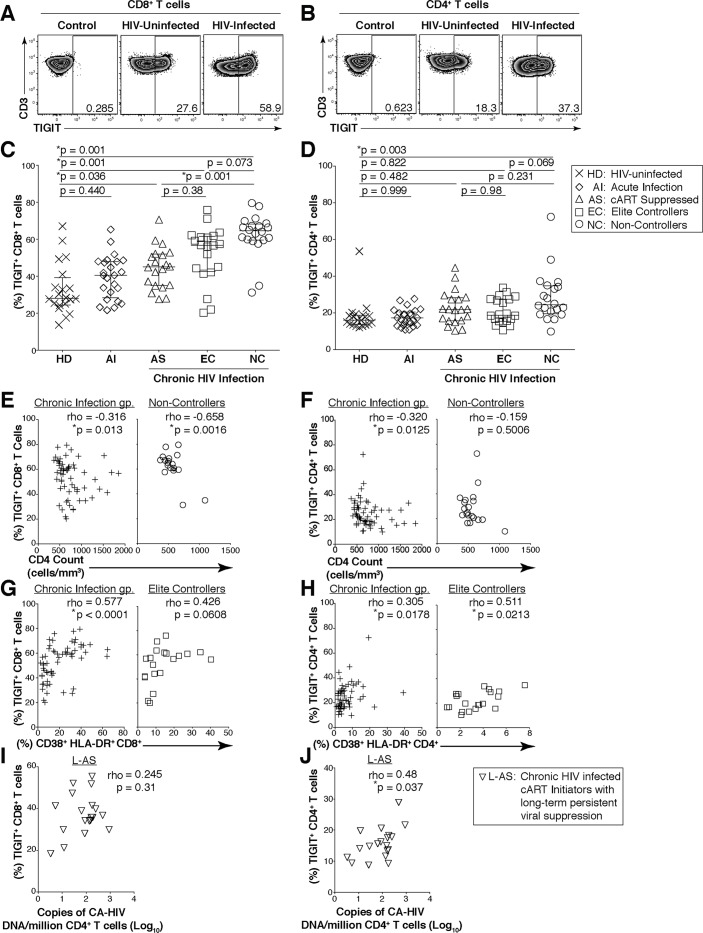
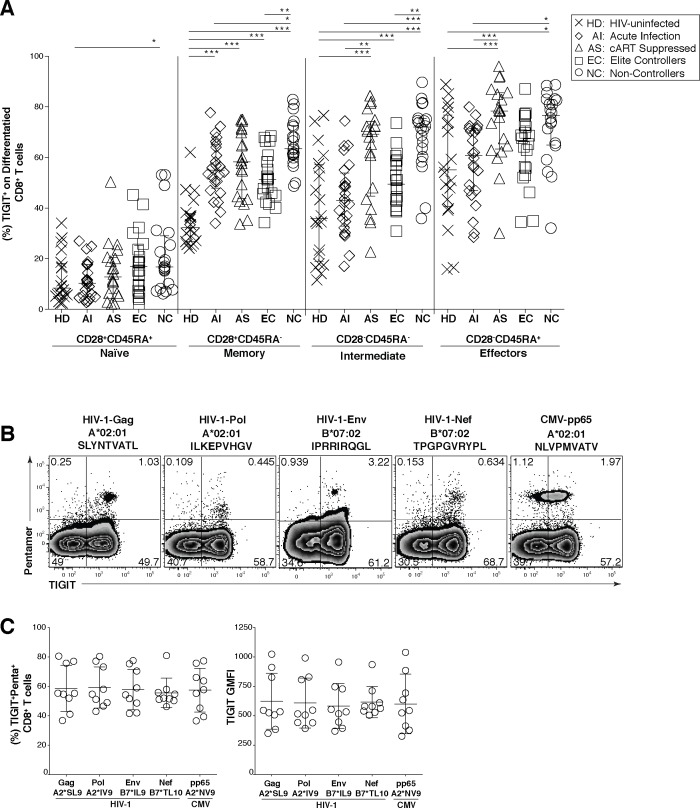
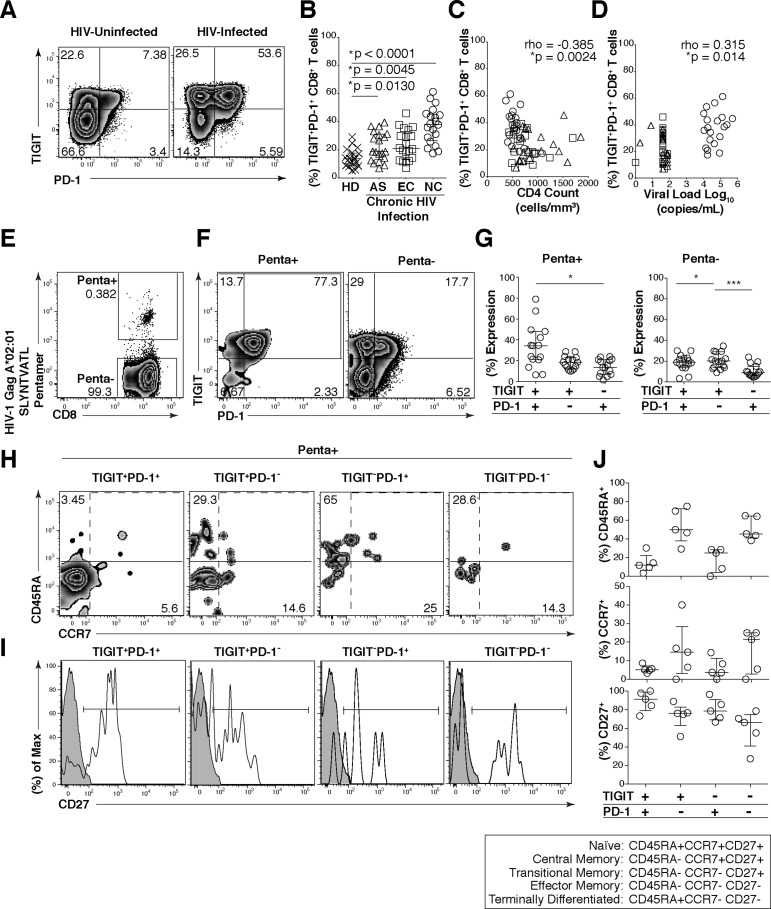
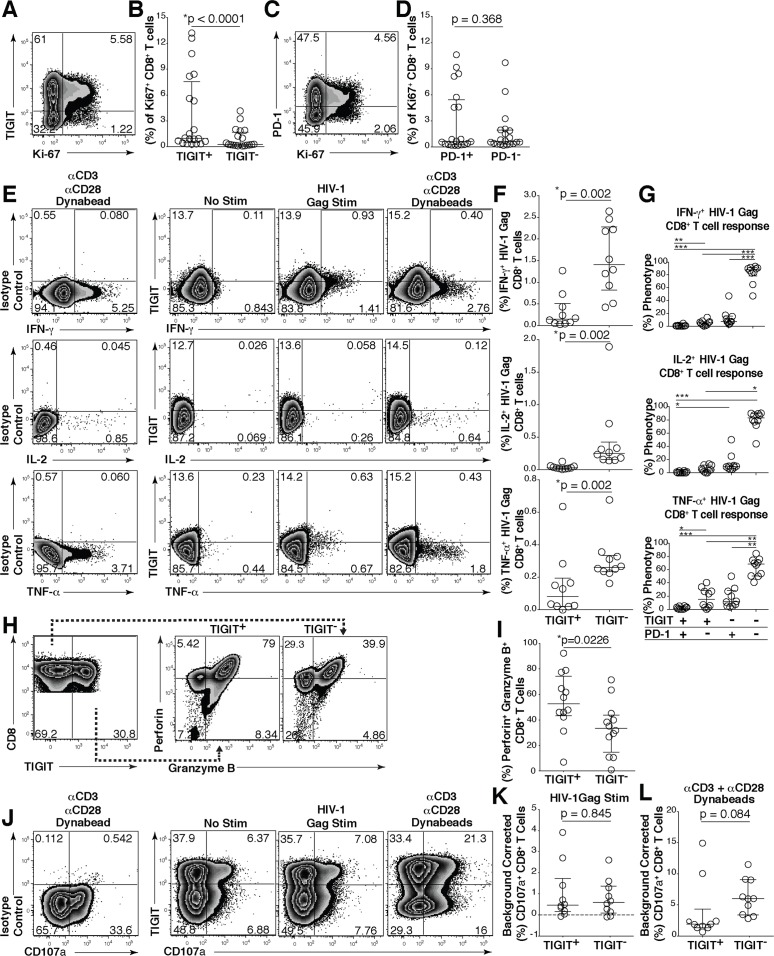
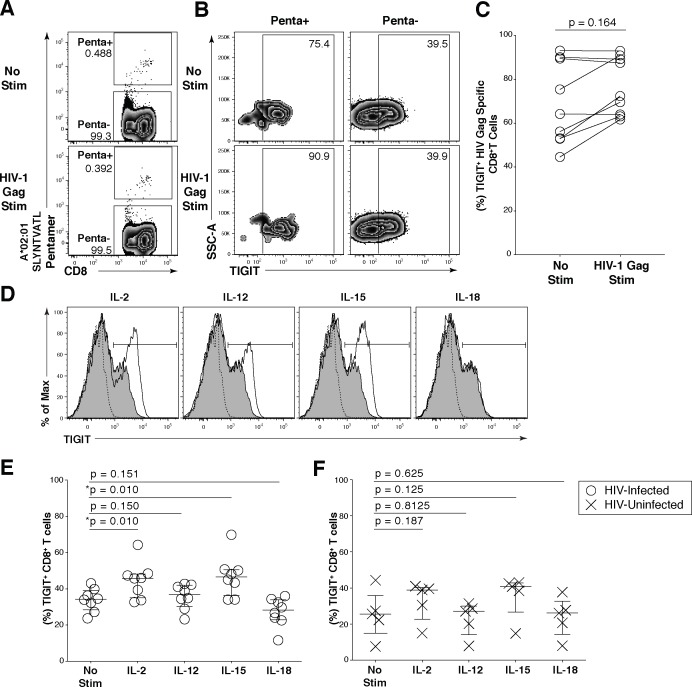
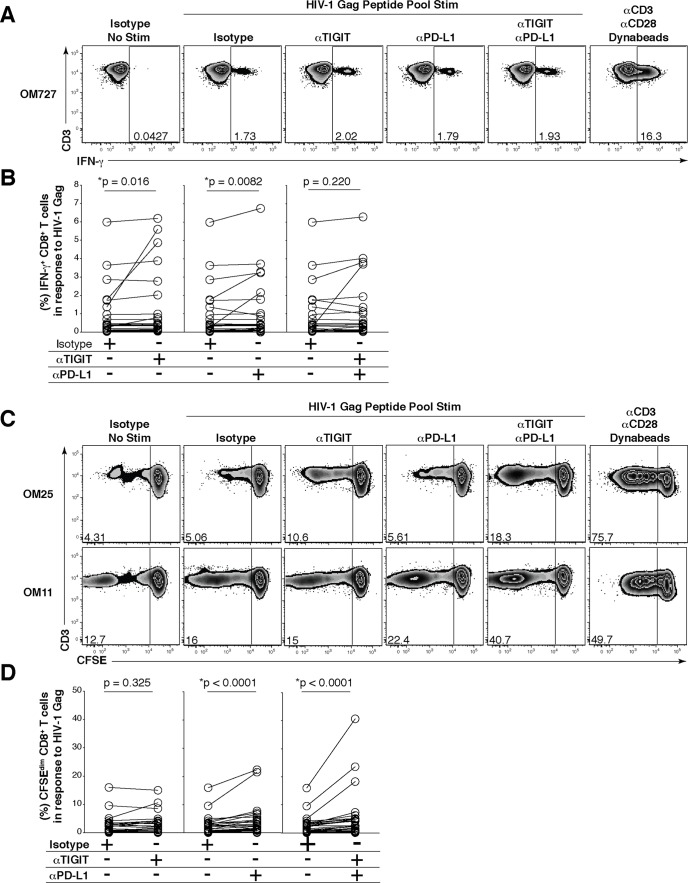
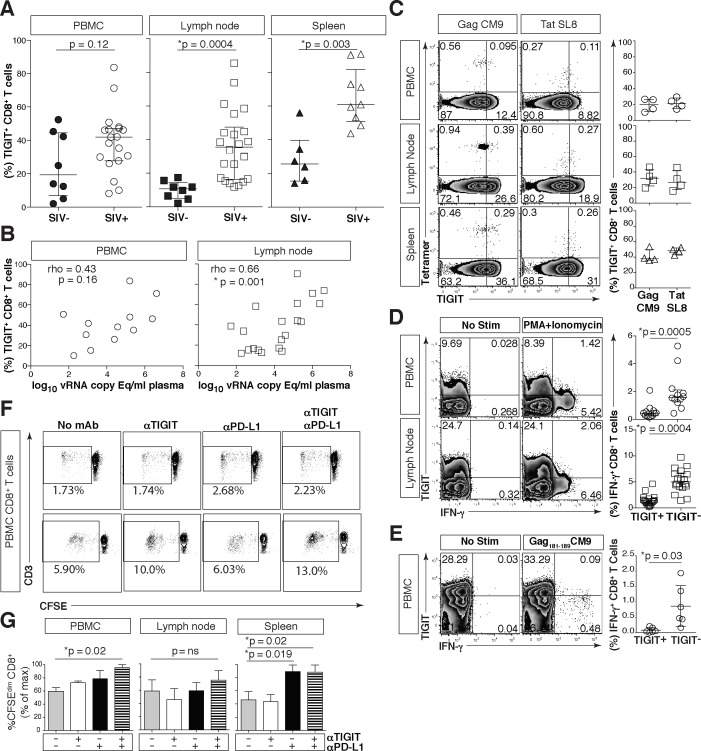
HIV infection induces phenotypic and functional changes to CD8+ T cells defined by the coordinated upregulation of a series of negative checkpoint receptors that eventually result in T cell exhaustion and failure to control viral replication. We report that effector CD8+ T cells during HIV infection in blood and SIV infection in lymphoid tissue exhibit higher levels of the negative checkpoint receptor TIGIT. Increased frequencies of TIGIT+ and TIGIT+ PD-1+ CD8+ T cells correlated with parameters of HIV and SIV disease progression. TIGIT remained elevated despite viral suppression in those with either pharmacological antiretroviral control or immunologically in elite controllers. HIV and SIV-specific CD8+ T cells were dysfunctional and expressed high levels of TIGIT and PD-1. Ex-vivo single or combinational antibody blockade of TIGIT and/or PD-L1 restored viral-specific CD8+ T cell effector responses. The frequency of TIGIT+ CD4+ T cells correlated with the CD4+ T cell total HIV DNA. These findings identify TIGIT as a novel marker of dysfunctional HIV-specific T cells and suggest TIGIT along with other checkpoint receptors may be novel curative HIV targets to reverse T cell exhaustion.
Conflict of interest statement
I hereby declare that MM (Scientist, Immuno-Oncology) and AJK (Vice president, Immuno-Oncology) are employed by Bristol-Myers Squibb, and have no significant competing financial, professional or personal interests that might have influenced the performance or presentation of the work described in this manuscript. This does not alter our adherence to all PLOS Pathogens policies on sharing data and materials.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
