

Phenotypic plasticity within yeast colonies: differential partitioning of cell fates
- PMID: 26743103
- PMCID: PMC4826809
- DOI: 10.1007/s00294-015-0558-y
Phenotypic plasticity within yeast colonies: differential partitioning of cell fates
Abstract
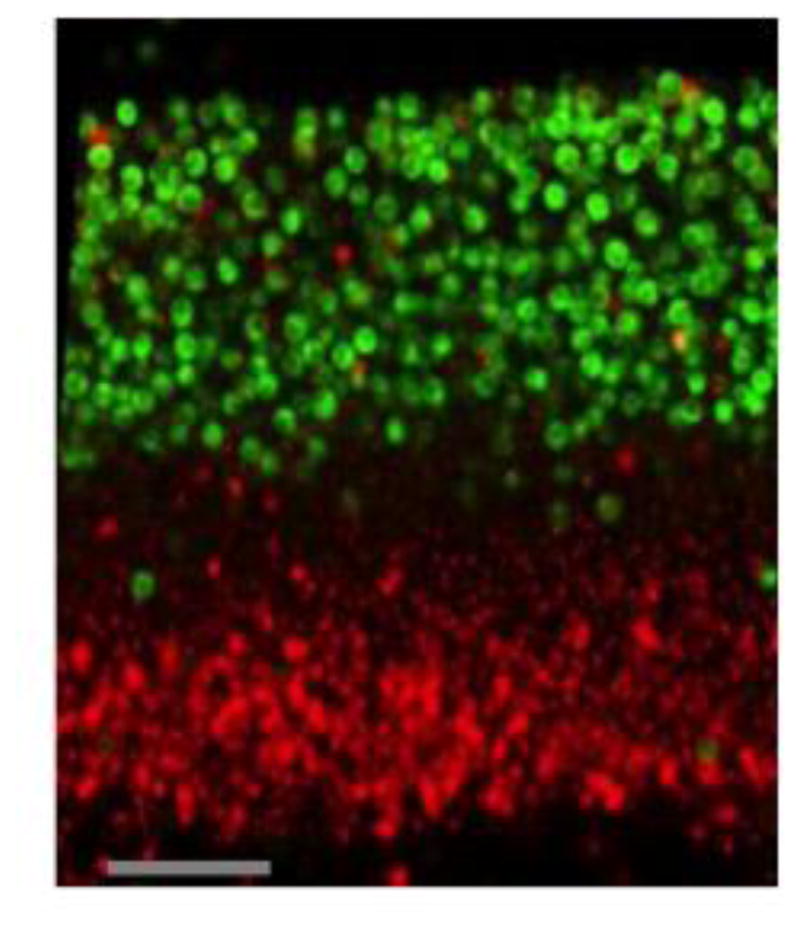
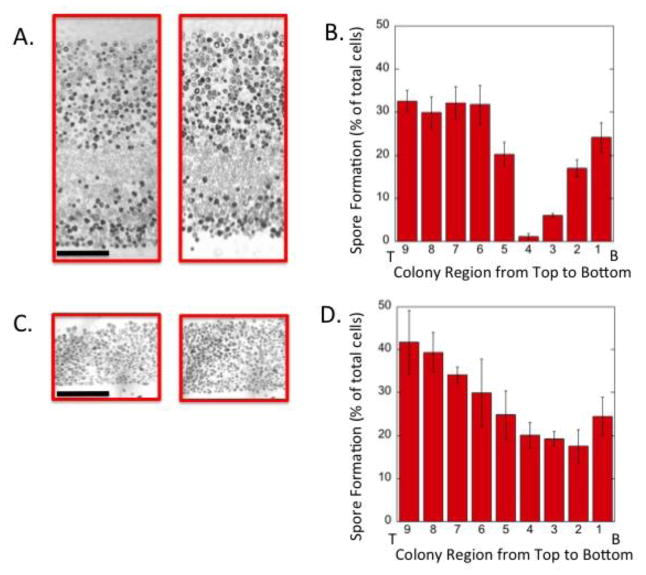
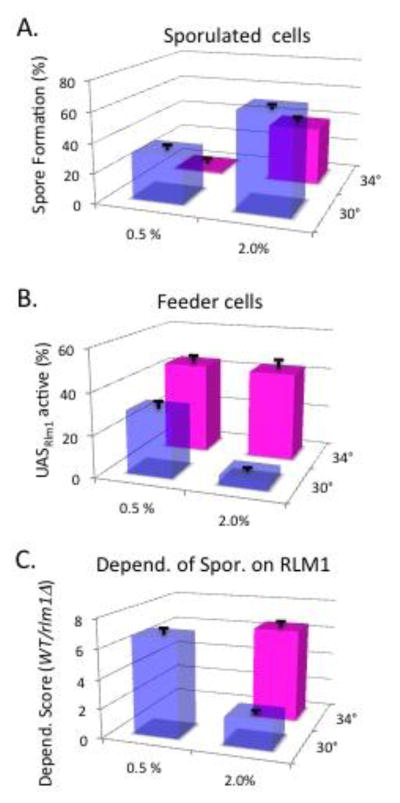
Across many phyla, a common aspect of multicellularity is the organization of different cell types into spatial patterns. In the budding yeast Saccharomyces cerevisiae, after diploid colonies have completed growth, they differentiate to form alternating layers of sporulating cells and feeder cells. In the current study, we found that as yeast colonies developed, the feeder cell layer was initially separated from the sporulating cell layer. Furthermore, the spatial pattern of sporulation in colonies depended on the colony's nutrient environment; in two environments in which overall colony sporulation efficiency was very similar, the pattern of feeder and sporulating cells within the colony was very different. As noted previously, under moderately suboptimal conditions for sporulation-low acetate concentration or high temperature-the number of feeder cells increases as does the dependence of sporulation on the feeder-cell transcription factor, Rlm1. Here we report that even under a condition that is completely blocked sporulation, the number of feeder cells still increased. These results suggest broader implications to our recently proposed "Differential Partitioning provides Environmental Buffering" or DPEB hypothesis.
Keywords: Allocation; Cell-wall integrity; IME2; Meiosis; Quiescent; RLM1.
Figures
Similar articles
-
Cell Differentiation and Spatial Organization in Yeast Colonies: Role of Cell-Wall Integrity Pathway.Genetics. 2015 Dec;201(4):1427-38. doi: 10.1534/genetics.115.180919. Epub 2015 Oct 28. Genetics. 2015. PMID: 26510787 Free PMC article.
-
How Boundaries Form: Linked Nonautonomous Feedback Loops Regulate Pattern Formation in Yeast Colonies.Genetics. 2019 Dec;213(4):1373-1386. doi: 10.1534/genetics.119.302700. Epub 2019 Oct 16. Genetics. 2019. PMID: 31619446 Free PMC article.
-
The Rim101p/PacC pathway and alkaline pH regulate pattern formation in yeast colonies.Genetics. 2010 Mar;184(3):707-16. doi: 10.1534/genetics.109.113480. Epub 2009 Dec 28. Genetics. 2010. PMID: 20038633 Free PMC article.
-
Sporulation in the budding yeast Saccharomyces cerevisiae.Genetics. 2011 Nov;189(3):737-65. doi: 10.1534/genetics.111.127126. Genetics. 2011. PMID: 22084423 Free PMC article. Review.
-
Cross-talk between autophagy and sporulation in Saccharomyces cerevisiae.Yeast. 2021 Jul;38(7):401-413. doi: 10.1002/yea.3556. Epub 2021 Mar 15. Yeast. 2021. PMID: 33608896 Review.
Cited by
-
Shrinking Daughters: Rlm1-Dependent G1/S Checkpoint Maintains Saccharomyces cerevisiae Daughter Cell Size and Viability.Genetics. 2017 Aug;206(4):1923-1938. doi: 10.1534/genetics.117.204206. Epub 2017 Jun 21. Genetics. 2017. PMID: 28637712 Free PMC article.
-
Mec1ATR is needed for extensive telomere elongation in response to ethanol in yeast.Curr Genet. 2018 Feb;64(1):223-234. doi: 10.1007/s00294-017-0728-1. Epub 2017 Aug 5. Curr Genet. 2018. PMID: 28780613
-
Sporulation: how to survive on planet Earth (and beyond).Curr Genet. 2017 Oct;63(5):831-838. doi: 10.1007/s00294-017-0694-7. Epub 2017 Apr 18. Curr Genet. 2017. PMID: 28421279 Free PMC article. Review.
-
Aspects of Multicellularity in Saccharomyces cerevisiae Yeast: A Review of Evolutionary and Physiological Mechanisms.Genes (Basel). 2020 Jun 24;11(6):690. doi: 10.3390/genes11060690. Genes (Basel). 2020. PMID: 32599749 Free PMC article. Review.
-
Cell differentiation, aging, and death in spatially organized yeast communities: mechanisms and consequences.Cell Death Differ. 2025 Mar 29. doi: 10.1038/s41418-025-01485-9. Online ahead of print. Cell Death Differ. 2025. PMID: 40158069 Review.
References
-
- Brakefield PM, Gems D, Cowen T, Christensen K, Grubeck-Loebenstein B, Keller L, … Westendorp RG. What are the effects of maternal and pre-adult environments on ageing in humans, and are there lessons from animal models? Mechanisms of ageing and development. 2005;126(3):431–438. doi: 10.1016/j.mad.2004.07.013. - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
