

Comparative Analysis of the Flax Immune Receptors L6 and L7 Suggests an Equilibrium-Based Switch Activation Model
- PMID: 26744216
- PMCID: PMC4746675
- DOI: 10.1105/tpc.15.00303
Comparative Analysis of the Flax Immune Receptors L6 and L7 Suggests an Equilibrium-Based Switch Activation Model
Abstract
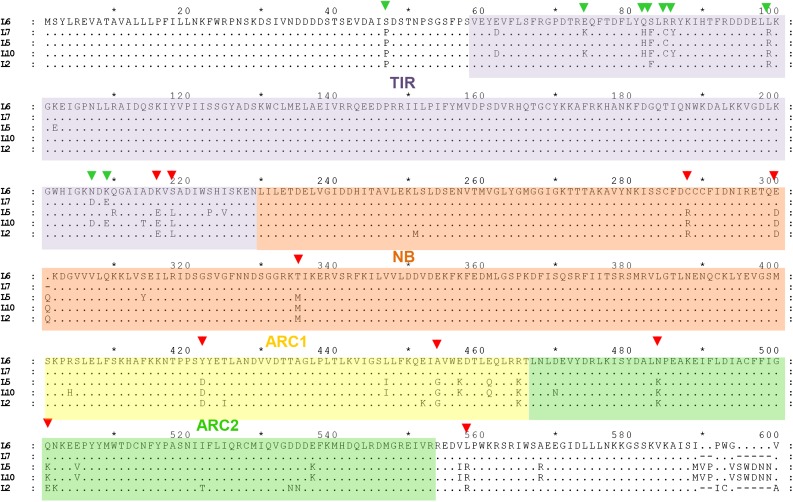
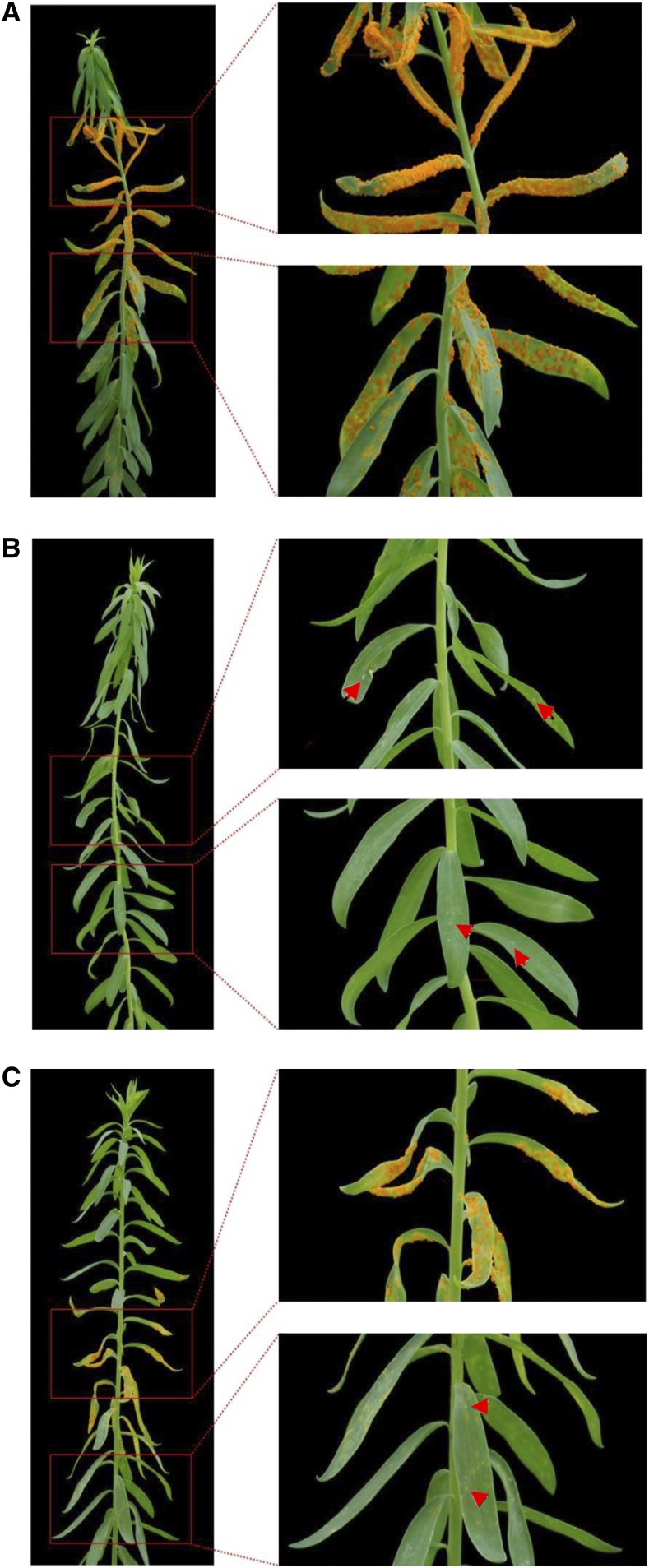
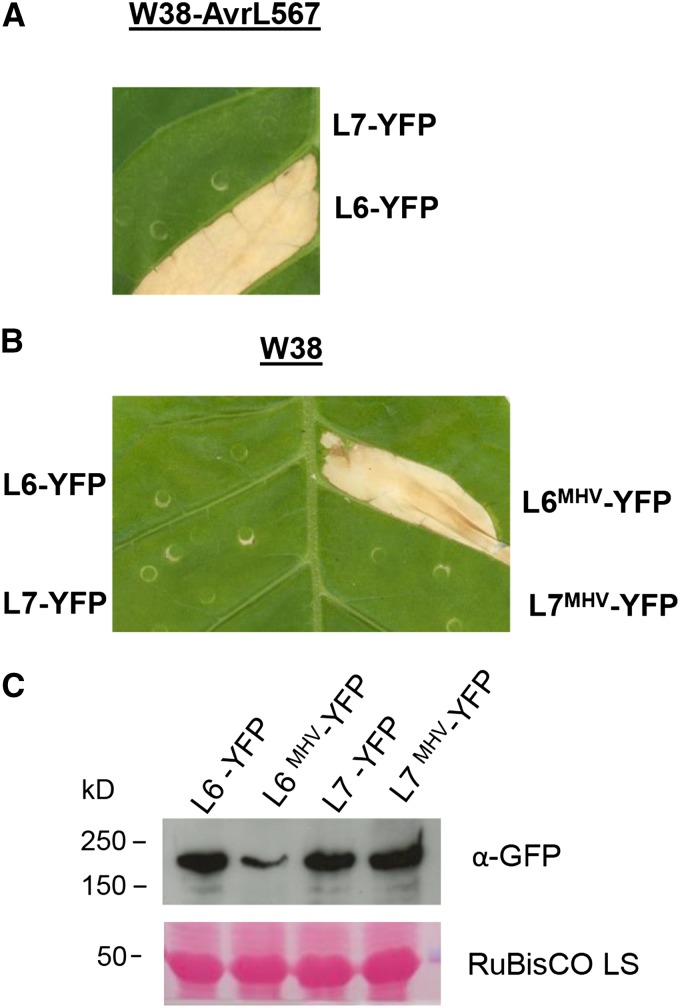
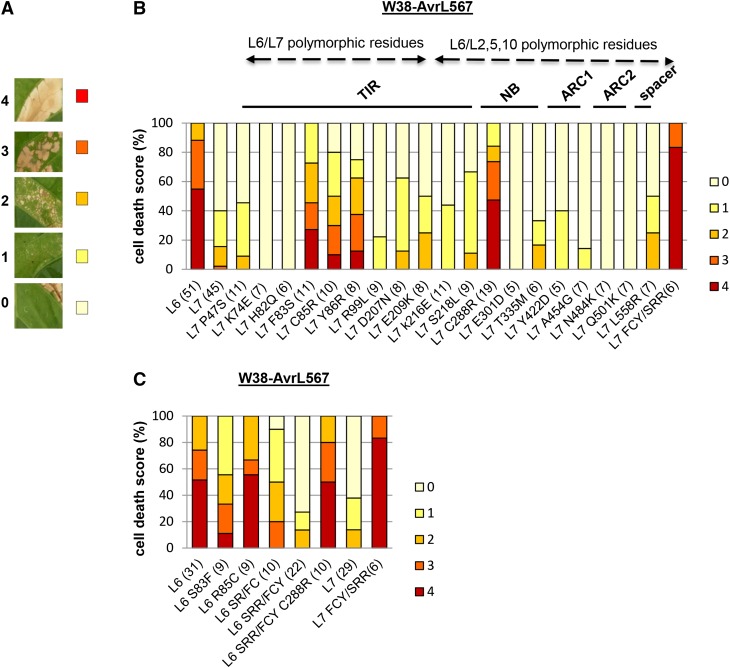
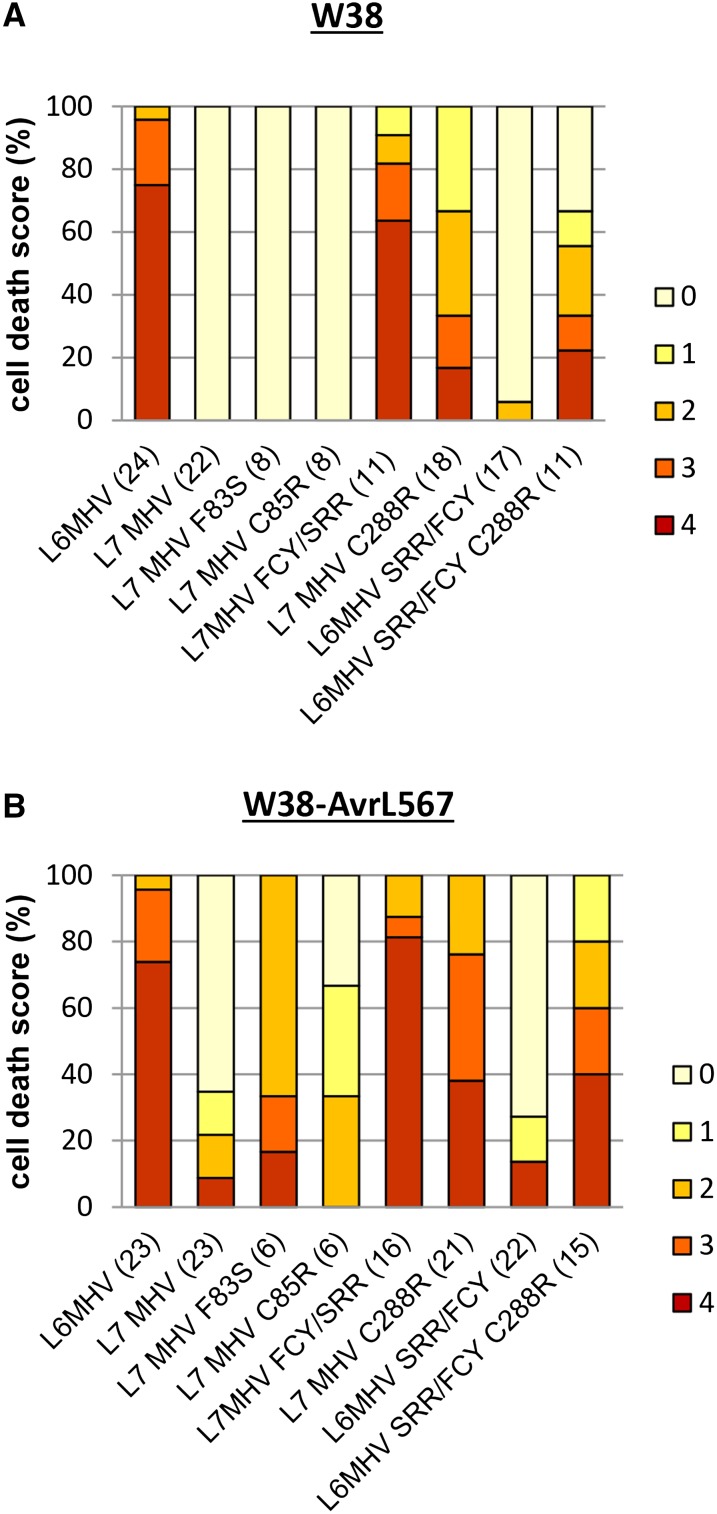
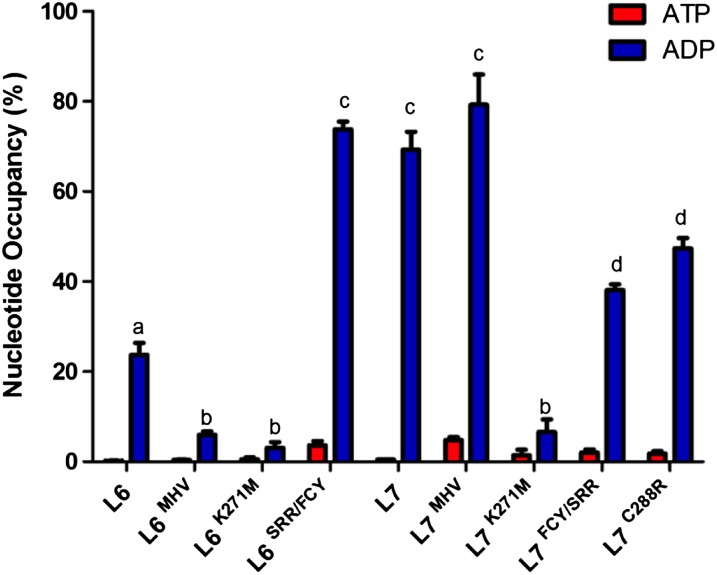
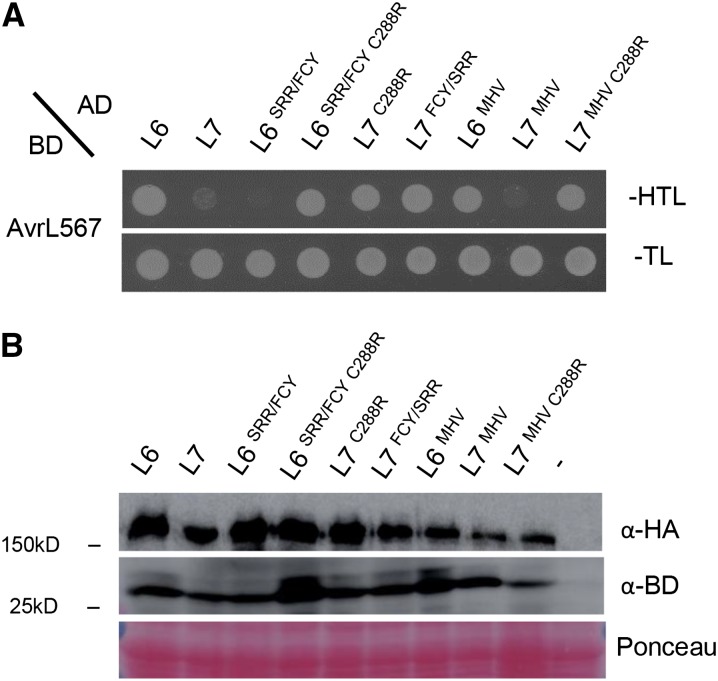
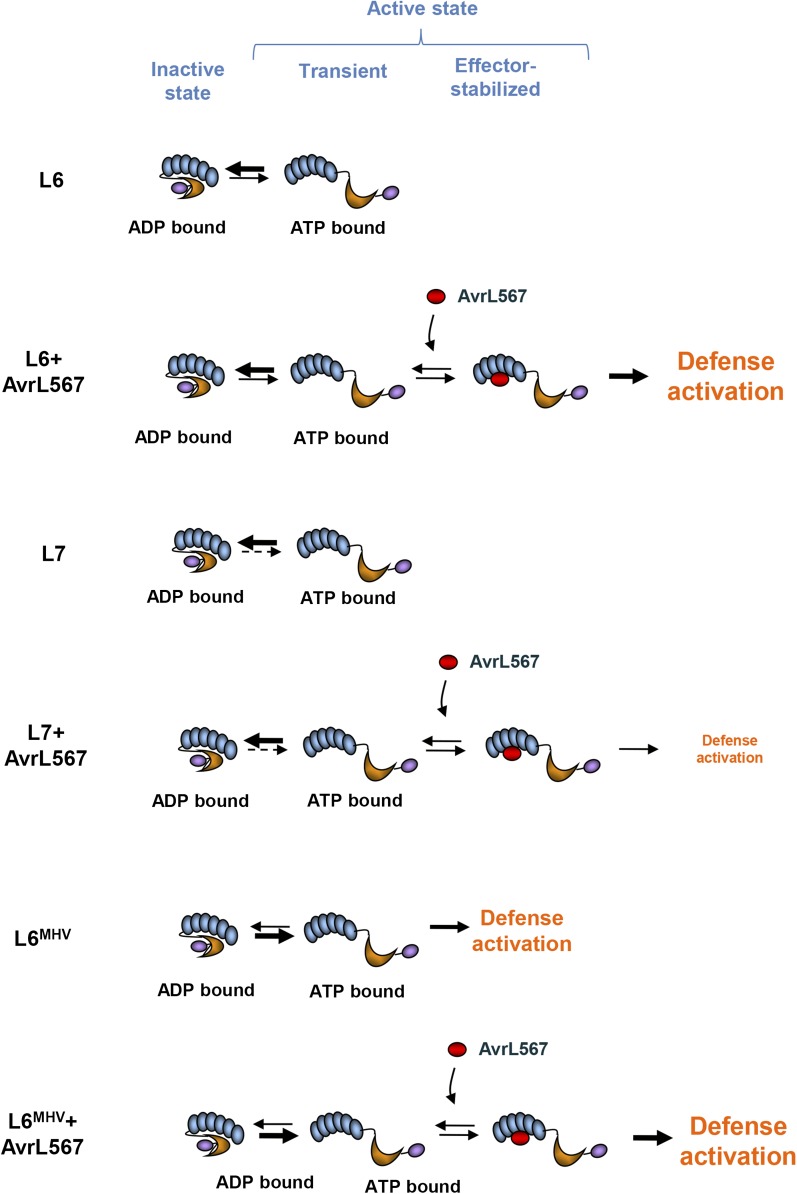
NOD-like receptors (NLRs) are central components of the plant immune system. L6 is a Toll/interleukin-1 receptor (TIR) domain-containing NLR from flax (Linum usitatissimum) conferring immunity to the flax rust fungus. Comparison of L6 to the weaker allele L7 identified two polymorphic regions in the TIR and the nucleotide binding (NB) domains that regulate both effector ligand-dependent and -independent cell death signaling as well as nucleotide binding to the receptor. This suggests that a negative functional interaction between the TIR and NB domains holds L7 in an inactive/ADP-bound state more tightly than L6, hence decreasing its capacity to adopt the active/ATP-bound state and explaining its weaker activity in planta. L6 and L7 variants with a more stable ADP-bound state failed to bind to AvrL567 in yeast two-hybrid assays, while binding was detected to the signaling active variants. This contrasts with current models predicting that effectors bind to inactive receptors to trigger activation. Based on the correlation between nucleotide binding, effector interaction, and immune signaling properties of L6/L7 variants, we propose that NLRs exist in an equilibrium between ON and OFF states and that effector binding to the ON state stabilizes this conformation, thereby shifting the equilibrium toward the active form of the receptor to trigger defense signaling.
© 2016 American Society of Plant Biologists. All rights reserved.
Figures
References
-
- Axtell M.J., Staskawicz B.J. (2003). Initiation of RPS2-specified disease resistance in Arabidopsis is coupled to the AvrRpt2-directed elimination of RIN4. Cell 112: 369–377. - PubMed
-
- Bao Q., Riedl S.J., Shi Y. (2005). Structure of Apaf-1 in the auto-inhibited form: a critical role for ADP. Cell Cycle 4: 1001–1003. - PubMed
-
- Bendahmane A., Farnham G., Moffett P., Baulcombe D.C. (2002). Constitutive gain-of-function mutants in a nucleotide binding site-leucine rich repeat protein encoded at the Rx locus of potato. Plant J. 32: 195–204. - PubMed
-
- Bent A.F., Kunkel B.N., Dahlbeck D., Brown K.L., Schmidt R., Giraudat J., Leung J., Staskawicz B.J. (1994). RPS2 of Arabidopsis thaliana: a leucine-rich repeat class of plant disease resistance genes. Science 265: 1856–1860. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
