

Accurate concentration control of mitochondria and nucleoids
- PMID: 26744405
- PMCID: PMC4823142
- DOI: 10.1126/science.aaa8714
Accurate concentration control of mitochondria and nucleoids
Abstract
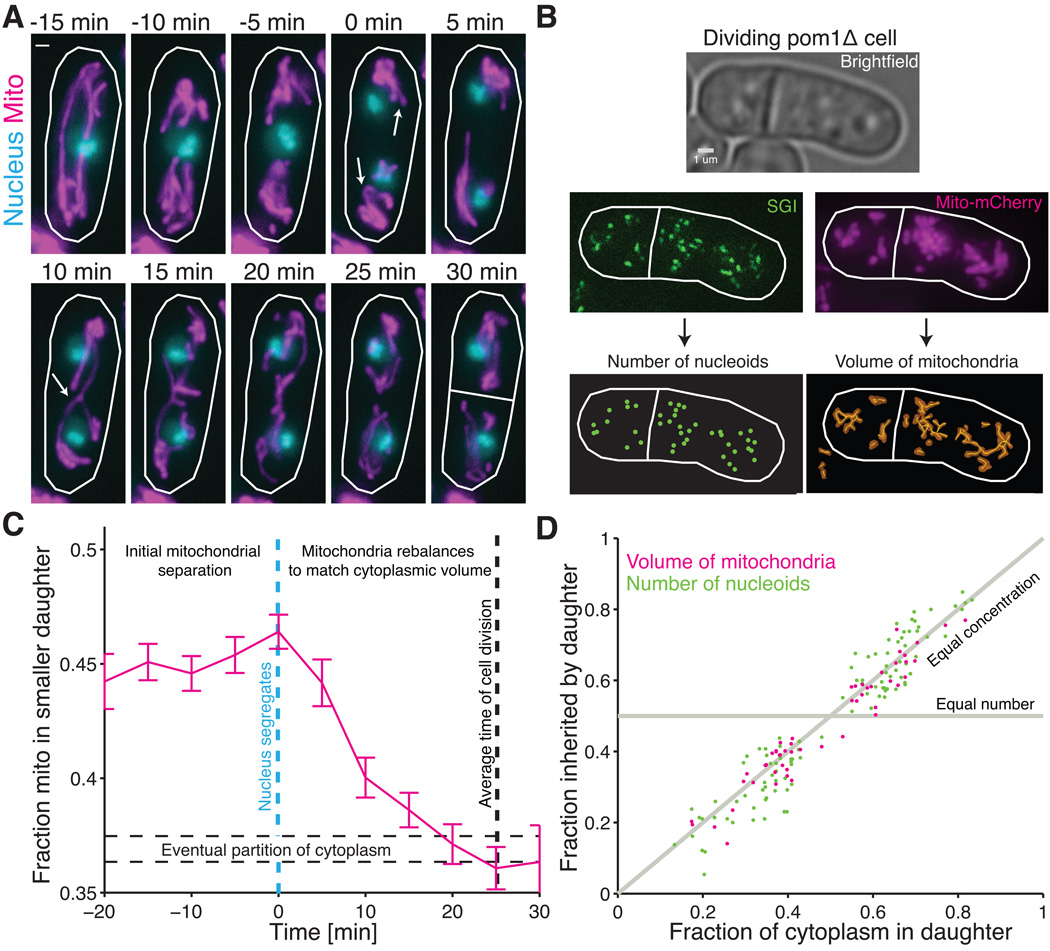
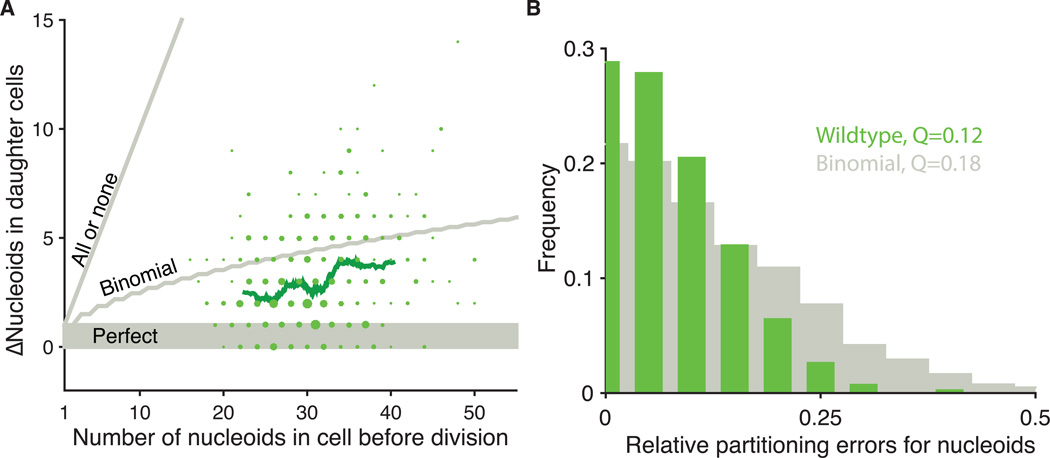
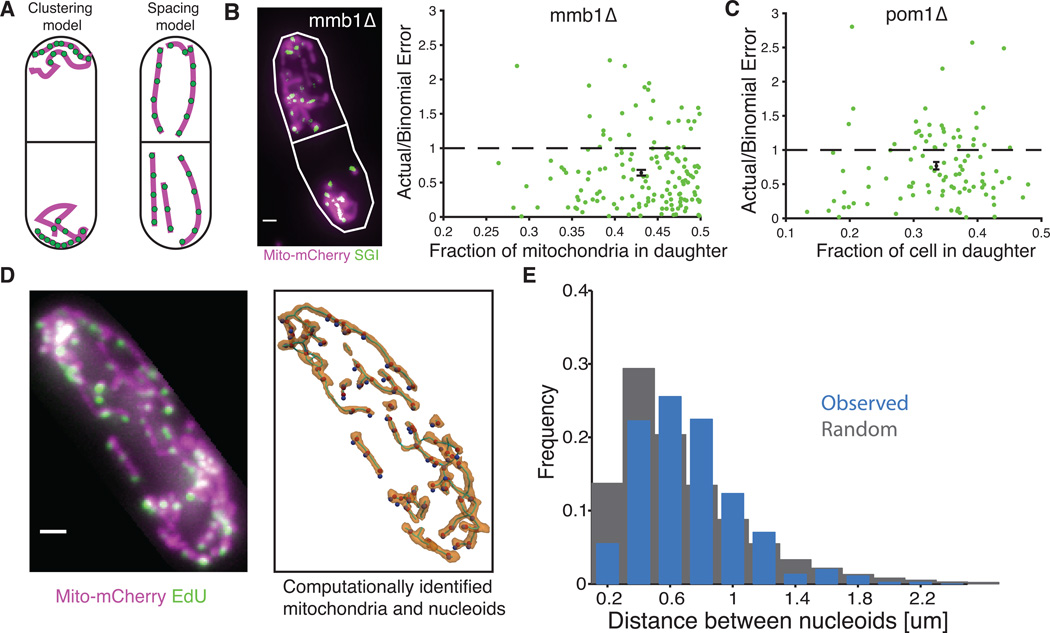
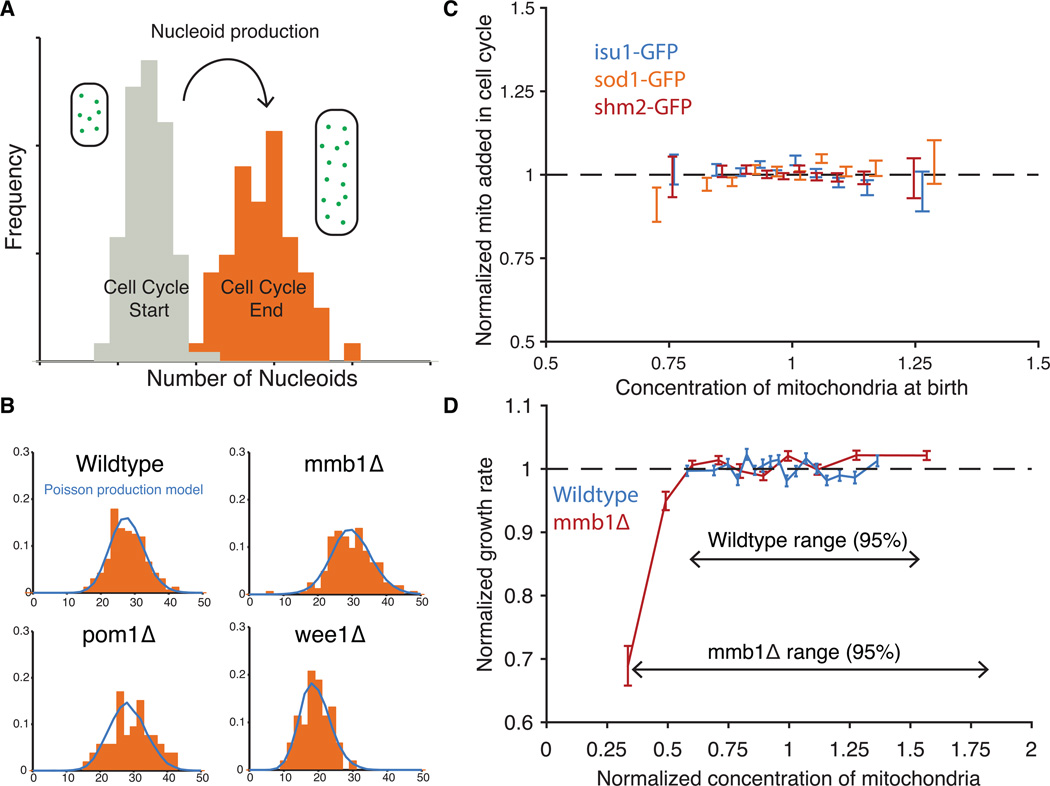
All cellular materials are partitioned between daughters at cell division, but by various mechanisms and with different accuracy. In the yeast Schizosaccharomyces pombe, the mitochondria are pushed to the cell poles by the spindle. We found that mitochondria spatially reequilibrate just before division, and that the mitochondrial volume and DNA-containing nucleoids instead segregate in proportion to the cytoplasm inherited by each daughter. However, nucleoid partitioning errors are suppressed by control at two levels: Mitochondrial volume is actively distributed throughout a cell, and nucleoids are spaced out in semiregular arrays within mitochondria. During the cell cycle, both mitochondria and nucleoids appear to be produced without feedback, creating a net control of fluctuations that is just accurate enough to avoid substantial growth defects.
Copyright © 2016, American Association for the Advancement of Science.
Figures
References
-
- Liu Z, Butow RA. Mitochondrial Retrograde Signaling. Annu. Rev. Genet. 2006;40:159–185. - PubMed
-
- Hoppins S, Nunnari J. Mitochondrial Dynamics and Apoptosis--the ER Connection. Science. 2012;337:1052–1054. - PubMed
-
- Duchen MR. Mitochondria in health and disease: perspectives on a new mitochondrial biology. Molecular Aspects of Medicine. 2004;25:365–451. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
