

Epigenetic regulation of cyclooxygenase-2 by methylation of c8orf4 in pulmonary fibrosis
- PMID: 26744410
- PMCID: PMC4782165
- DOI: 10.1042/CS20150697
Epigenetic regulation of cyclooxygenase-2 by methylation of c8orf4 in pulmonary fibrosis
Abstract
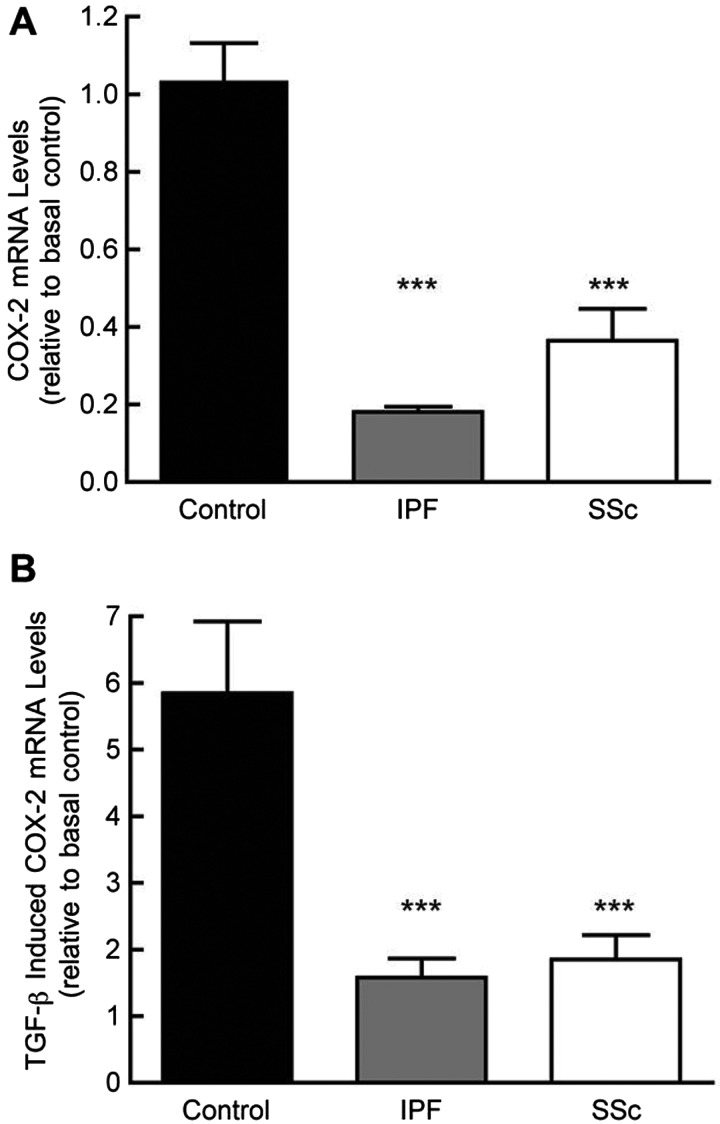
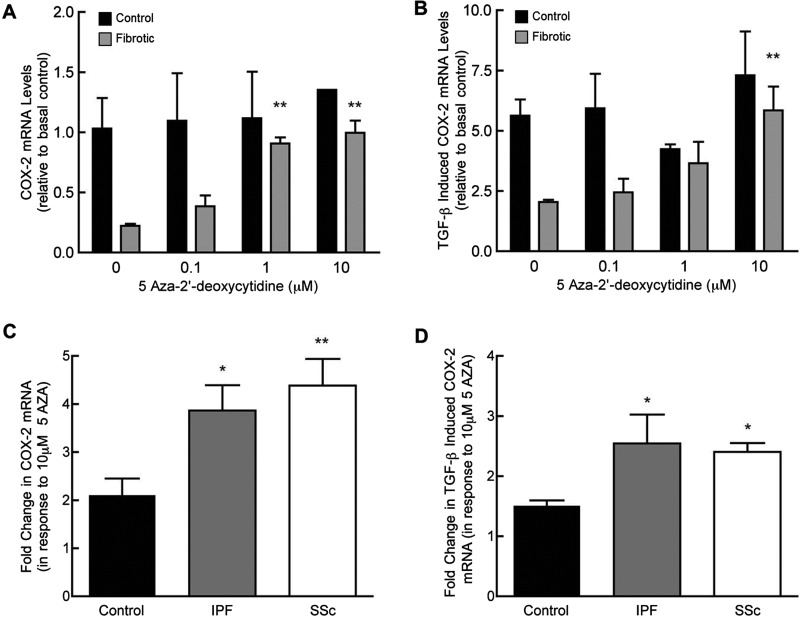
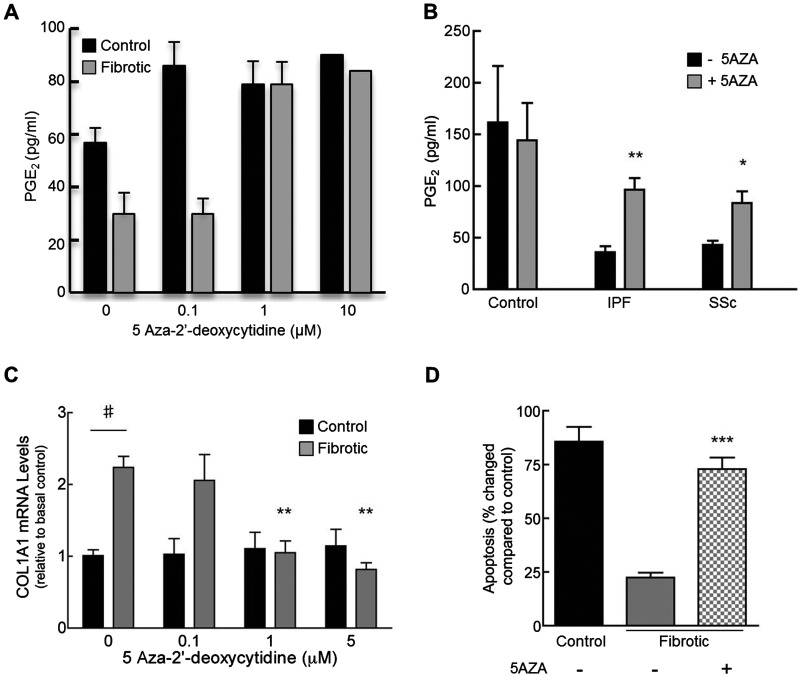
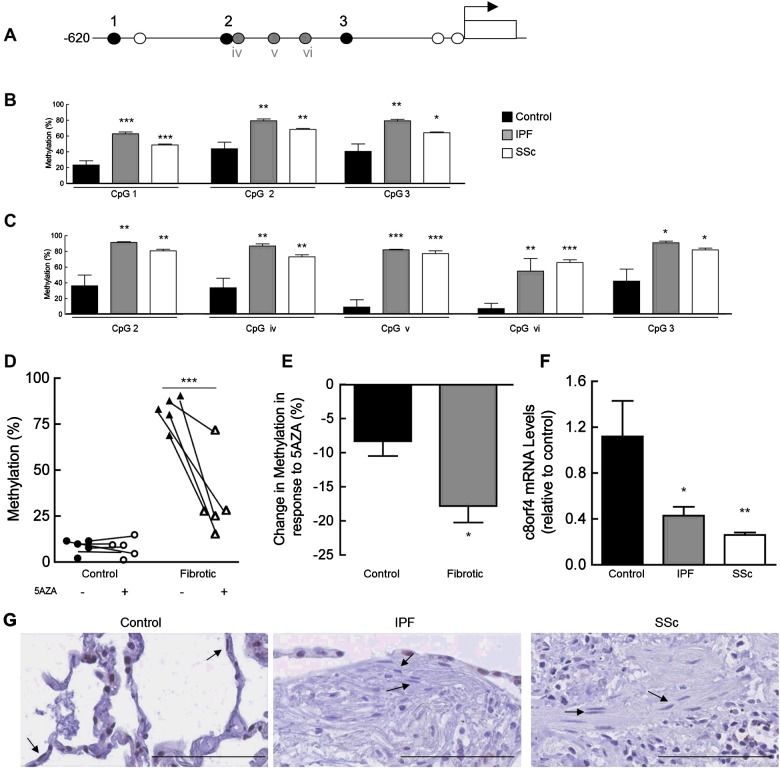
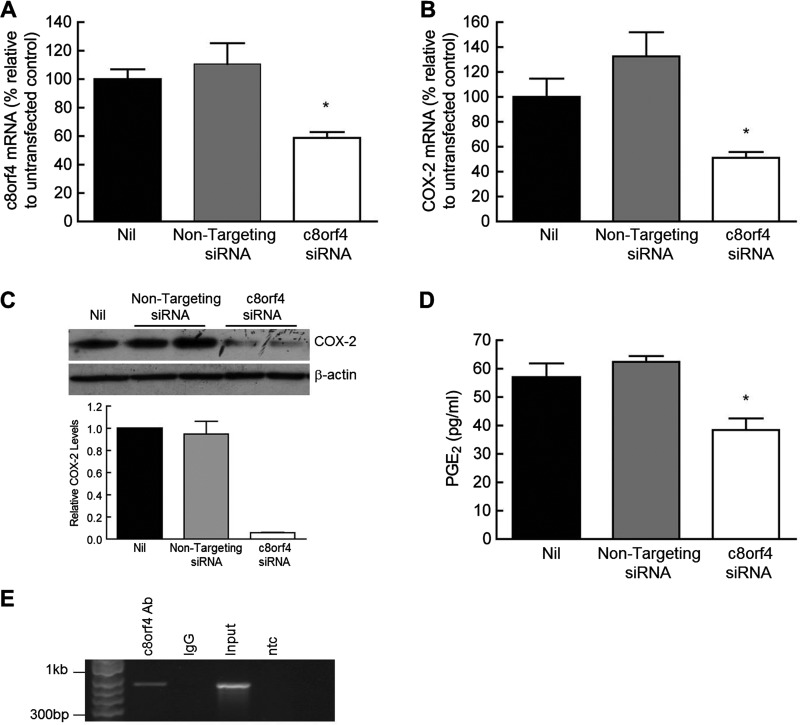
Fibroblasts derived from the lungs of patients with idiopathic pulmonary fibrosis (IPF) and systemic sclerosis (SSc) produce low levels of prostaglandin (PG) E2, due to a limited capacity to up-regulate cyclooxygenase-2 (COX-2). This deficiency contributes functionally to the fibroproliferative state, however the mechanisms responsible are incompletely understood. In the present study, we examined whether the reduced level of COX-2 mRNA expression observed in fibrotic lung fibroblasts is regulated epigenetically. The DNA methylation inhibitor, 5-aza-2'-deoxycytidine (5AZA) restored COX-2 mRNA expression by fibrotic lung fibroblasts dose dependently. Functionally, this resulted in normalization of fibroblast phenotype in terms of PGE2 production, collagen mRNA expression and sensitivity to apoptosis. COX-2 methylation assessed by bisulfite sequencing and methylation microarrays was not different in fibrotic fibroblasts compared with controls. However, further analysis of the methylation array data identified a transcriptional regulator, chromosome 8 open reading frame 4 (thyroid cancer protein 1, TC-1) (c8orf4), which is hypermethylated and down-regulated in fibrotic fibroblasts compared with controls. siRNA knockdown of c8orf4 in control fibroblasts down-regulated COX-2 and PGE2 production generating a phenotype similar to that observed in fibrotic lung fibroblasts. Chromatin immunoprecipitation demonstrated that c8orf4 regulates COX-2 expression in lung fibroblasts through binding of the proximal promoter. We conclude that the decreased capacity of fibrotic lung fibroblasts to up-regulate COX-2 expression and COX-2-derived PGE2 synthesis is due to an indirect epigenetic mechanism involving hypermethylation of the transcriptional regulator, c8orf4.
Keywords: DNA methylation; cyclooxygenase-2; fibroblast; idiopathic pulmonary fibrosis; prostaglandin E2; systemic sclerosis.
© 2016 The Author(s).
Figures
Similar articles
-
Hypoxia-induced DNA hypermethylation in human pulmonary fibroblasts is associated with Thy-1 promoter methylation and the development of a pro-fibrotic phenotype.Respir Res. 2012 Aug 31;13(1):74. doi: 10.1186/1465-9921-13-74. Respir Res. 2012. PMID: 22938014 Free PMC article.
-
Hypermethylation of PTGER2 confers prostaglandin E2 resistance in fibrotic fibroblasts from humans and mice.Am J Pathol. 2010 Nov;177(5):2245-55. doi: 10.2353/ajpath.2010.100446. Epub 2010 Oct 1. Am J Pathol. 2010. PMID: 20889571 Free PMC article.
-
A central role for G9a and EZH2 in the epigenetic silencing of cyclooxygenase-2 in idiopathic pulmonary fibrosis.FASEB J. 2014 Jul;28(7):3183-96. doi: 10.1096/fj.13-241760. Epub 2014 Mar 20. FASEB J. 2014. PMID: 24652950 Free PMC article.
-
Interplay between genetics and epigenetics in lung fibrosis.Int J Biochem Cell Biol. 2025 Mar;180:106739. doi: 10.1016/j.biocel.2025.106739. Epub 2025 Jan 21. Int J Biochem Cell Biol. 2025. PMID: 39848439 Review.
-
DNA Methylation of Fibroblast Phenotypes and Contributions to Lung Fibrosis.Cells. 2021 Aug 3;10(8):1977. doi: 10.3390/cells10081977. Cells. 2021. PMID: 34440746 Free PMC article. Review.
Cited by
-
More than a Genetic Code: Epigenetics of Lung Fibrosis.Mol Diagn Ther. 2020 Dec;24(6):665-681. doi: 10.1007/s40291-020-00490-7. Mol Diagn Ther. 2020. PMID: 32926347 Free PMC article. Review.
-
Epigenetics of scleroderma: Integrating genetic, ethnic, age, and environmental effects.J Scleroderma Relat Disord. 2019 Oct;4(3):238-250. doi: 10.1177/2397198319855872. Epub 2019 Jul 3. J Scleroderma Relat Disord. 2019. PMID: 35382507 Free PMC article. Review.
-
Adaptive and innate immune mechanisms in cardiac fibrosis complicating pulmonary arterial hypertension.Physiol Rep. 2020 Aug;8(15):e14532. doi: 10.14814/phy2.14532. Physiol Rep. 2020. PMID: 32786064 Free PMC article. Review.
-
DNA methylation in human diseases.Heliyon. 2024 Jun 4;10(11):e32366. doi: 10.1016/j.heliyon.2024.e32366. eCollection 2024 Jun 15. Heliyon. 2024. PMID: 38933971 Free PMC article. Review.
-
The Roles of Various Prostaglandins in Fibrosis: A Review.Biomolecules. 2021 May 24;11(6):789. doi: 10.3390/biom11060789. Biomolecules. 2021. PMID: 34073892 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials