

Trypanosoma cruzi Experimental Infection Impacts on the Thymic Regulatory T Cell Compartment
- PMID: 26745276
- PMCID: PMC4706328
- DOI: 10.1371/journal.pntd.0004285
Trypanosoma cruzi Experimental Infection Impacts on the Thymic Regulatory T Cell Compartment
Abstract
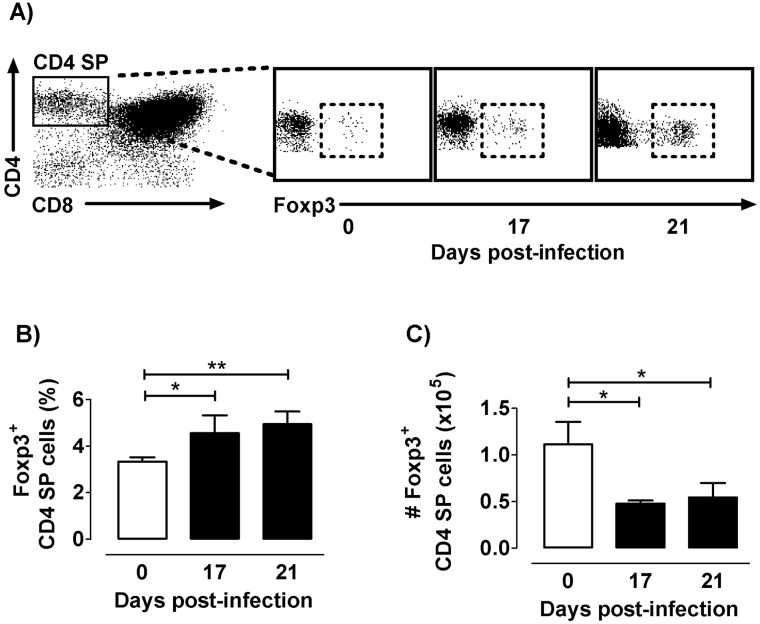
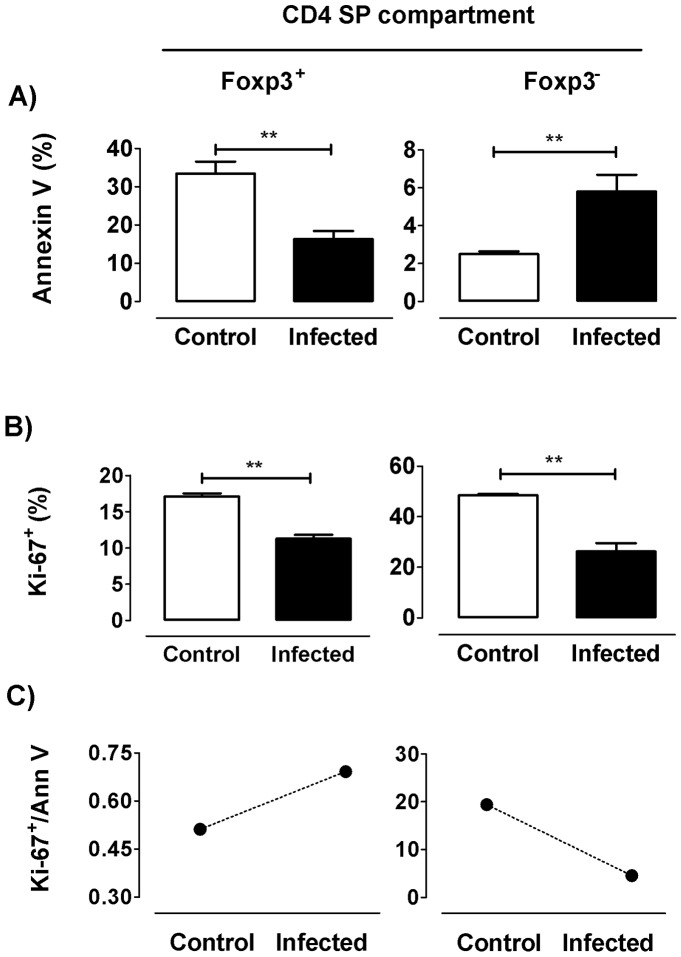
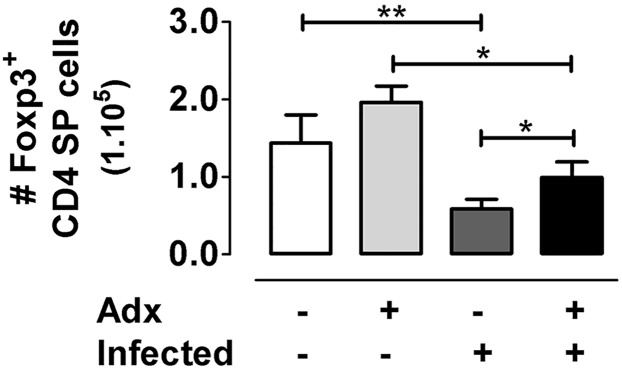
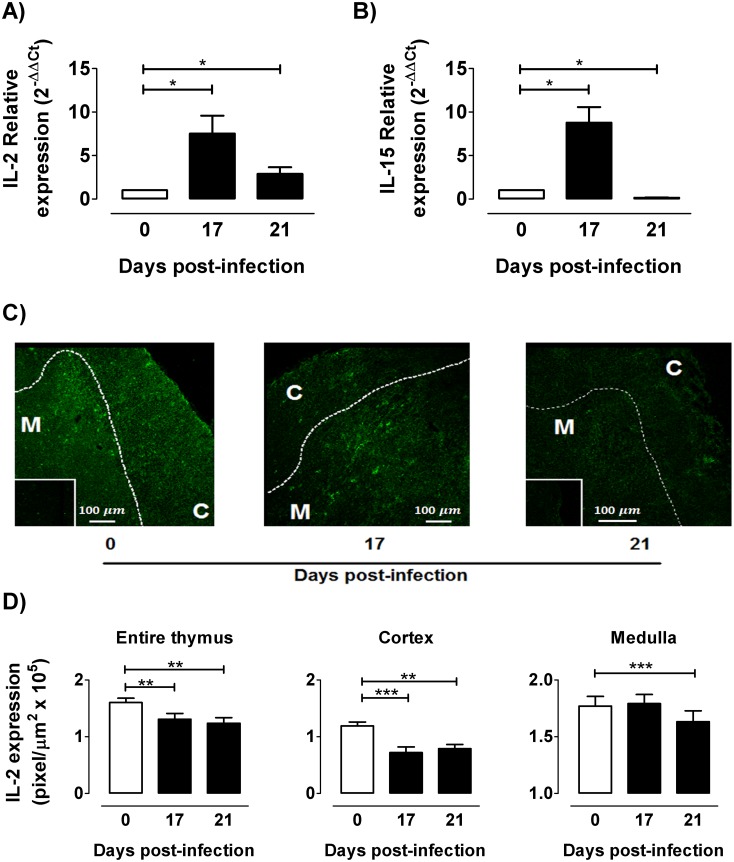
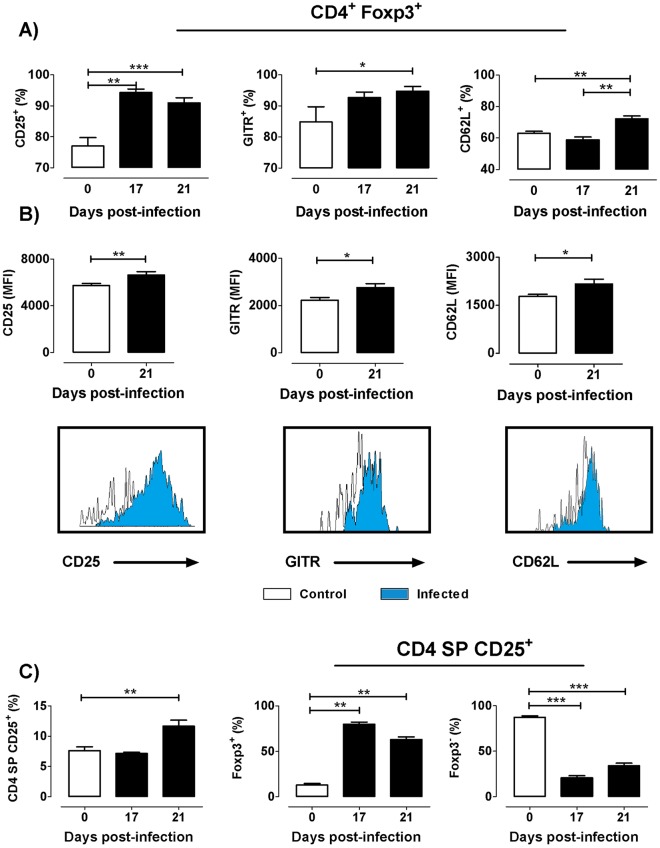
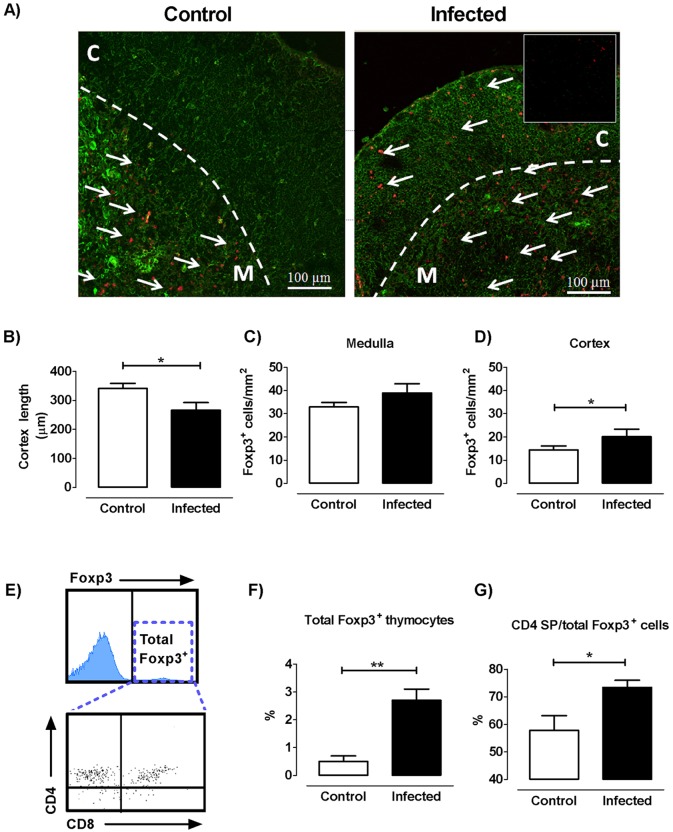
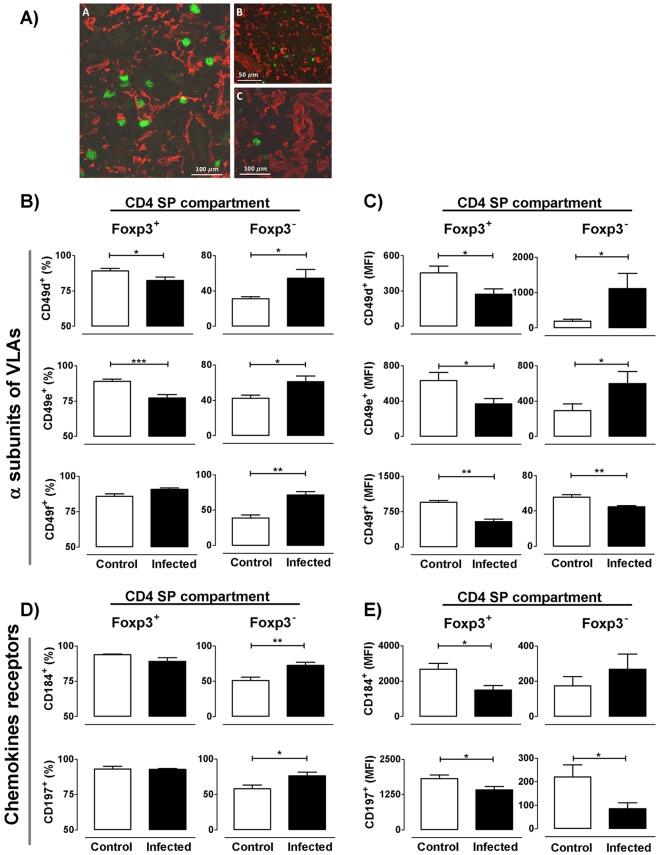
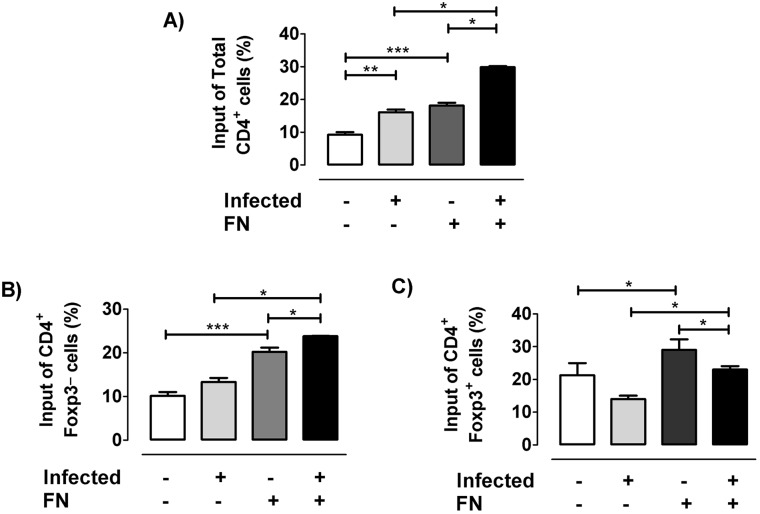
The dynamics of regulatory T cells in the course of Trypanosoma cruzi infection is still debated. We previously demonstrated that acute murine T. cruzi infection results in an impaired peripheral CD4+Foxp3+ T cell differentiation due to the acquisition of an abnormal Th1-like phenotype and altered functional features, negatively impacting on the course of infection. Moreover, T. cruzi infection induces an intense thymic atrophy. As known, the thymus is the primary lymphoid organ in which thymic-derived regulatory T cells, known as tTregs, differentiate. Considering the lack of available data about the effect of T. cruzi infection upon tTregs, we examined tTreg dynamics during the course of disease. We confirmed that T. cruzi infection induces a marked loss of tTreg cell number associated to cell precursor exhaustion, partially avoided by glucocorticoid ablation- and IL-2 survival factor depletion. At the same time, tTregs accumulate within the CD4 single-positive compartment, exhibiting an increased Ki-67/Annexin V ratio compared to controls. Moreover, tTregs enhance after the infection the expression of signature markers (CD25, CD62L and GITR) and they also display alterations in the expression of migration-associated molecules (α chains of VLAs and chemokine receptors) such as functional fibronectin-driven migratory disturbance. Taken together, we provide data demonstrating profound alterations in tTreg compartment during acute murine T. cruzi infection, denoting that their homeostasis is significantly affected. The evident loss of tTreg cell number may compromise the composition of tTreg peripheral pool, and such sustained alteration over time may be partially related to the immune dysregulation observed in the chronic phase of the disease.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Chagasic thymic atrophy does not affect negative selection but results in the export of activated CD4+CD8+ T cells in severe forms of human disease.PLoS Negl Trop Dis. 2011 Aug;5(8):e1268. doi: 10.1371/journal.pntd.0001268. Epub 2011 Aug 16. PLoS Negl Trop Dis. 2011. PMID: 21858238 Free PMC article.
-
Benznidazole therapy in Trypanosoma cruzi-infected mice blocks thymic involution and apoptosis of CD4+CD8+ double-positive thymocytes.Antimicrob Agents Chemother. 2005 May;49(5):1981-7. doi: 10.1128/AAC.49.5.1981-1987.2005. Antimicrob Agents Chemother. 2005. PMID: 15855522 Free PMC article.
-
Limited Foxp3+ Regulatory T Cells Response During Acute Trypanosoma cruzi Infection Is Required to Allow the Emergence of Robust Parasite-Specific CD8+ T Cell Immunity.Front Immunol. 2018 Nov 5;9:2555. doi: 10.3389/fimmu.2018.02555. eCollection 2018. Front Immunol. 2018. PMID: 30455700 Free PMC article.
-
Modulation of Intrathymic Sphingosine-1-Phosphate Levels Promotes Escape of Immature Thymocytes to the Periphery with a Potential Proinflammatory Role in Chagas Disease.Biomed Res Int. 2015;2015:709846. doi: 10.1155/2015/709846. Epub 2015 Aug 4. Biomed Res Int. 2015. PMID: 26347020 Free PMC article. Review.
-
[Role of cytokines in resistance and pathology in Trypanosoma cruzi infection].Rev Argent Microbiol. 1996 Apr-Jun;28(2):99-109. Rev Argent Microbiol. 1996. PMID: 8768488 Review. Spanish.
Cited by
-
MicroRNAs: master regulators in host-parasitic protist interactions.Open Biol. 2022 Jun;12(6):210395. doi: 10.1098/rsob.210395. Epub 2022 Jun 15. Open Biol. 2022. PMID: 35702995 Free PMC article. Review.
-
Role of Hormonal Circuitry Upon T Cell Development in Chagas Disease: Possible Implications on T Cell Dysfunctions.Front Endocrinol (Lausanne). 2018 Jun 14;9:334. doi: 10.3389/fendo.2018.00334. eCollection 2018. Front Endocrinol (Lausanne). 2018. PMID: 29963015 Free PMC article. Review.
-
Infection-Associated Thymic Atrophy.Front Immunol. 2021 May 25;12:652538. doi: 10.3389/fimmu.2021.652538. eCollection 2021. Front Immunol. 2021. PMID: 34113341 Free PMC article. Review.
-
Regulatory Lymphoid and Myeloid Cells Determine the Cardiac Immunopathogenesis of Trypanosoma cruzi Infection.Front Microbiol. 2018 Mar 1;9:351. doi: 10.3389/fmicb.2018.00351. eCollection 2018. Front Microbiol. 2018. PMID: 29545782 Free PMC article. Review.
-
Thymic Microenvironment Is Modified by Malnutrition and Leishmania infantum Infection.Front Cell Infect Microbiol. 2019 Jul 12;9:252. doi: 10.3389/fcimb.2019.00252. eCollection 2019. Front Cell Infect Microbiol. 2019. PMID: 31355153 Free PMC article.
References
-
- Belkaid Y. Regulatory T cells and infection: a dangerous necessity. Nat Rev Immunol. 2007;7: 875–888. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
