

A Higher Activation Threshold of Memory CD8+ T Cells Has a Fitness Cost That Is Modified by TCR Affinity during Tuberculosis
- PMID: 26745507
- PMCID: PMC4706326
- DOI: 10.1371/journal.ppat.1005380
A Higher Activation Threshold of Memory CD8+ T Cells Has a Fitness Cost That Is Modified by TCR Affinity during Tuberculosis
Abstract
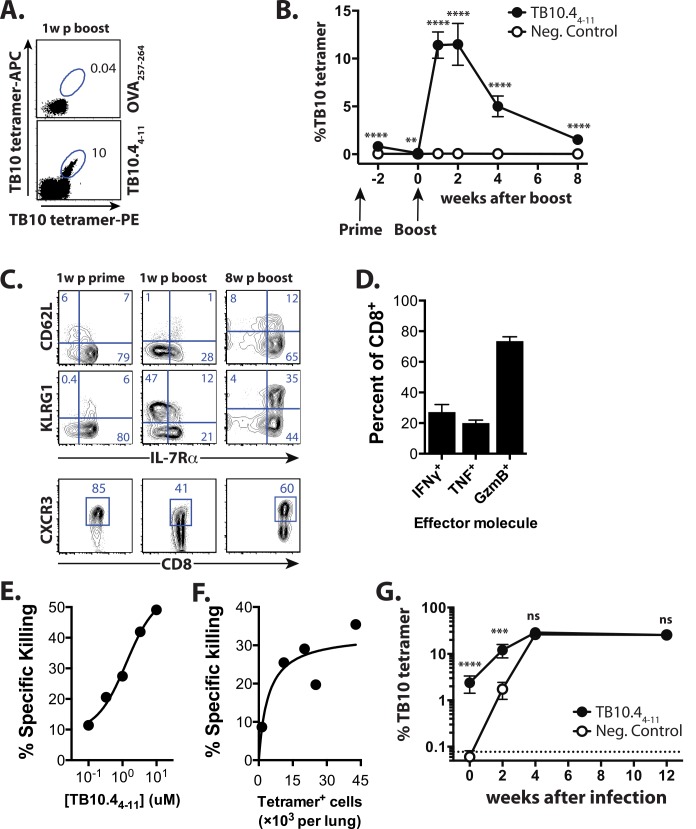
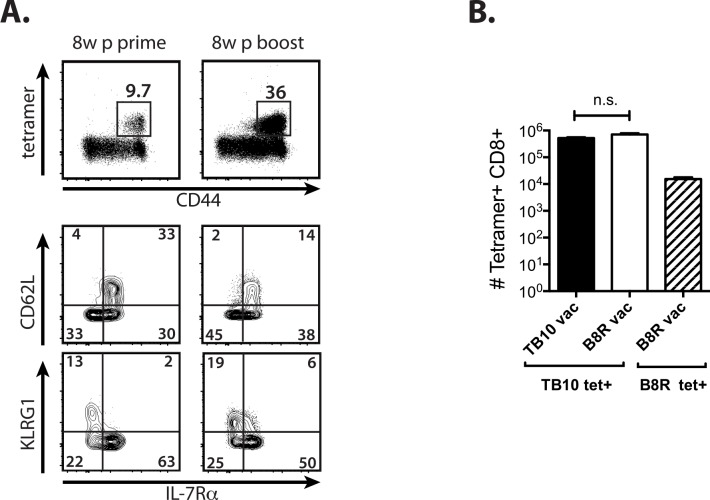
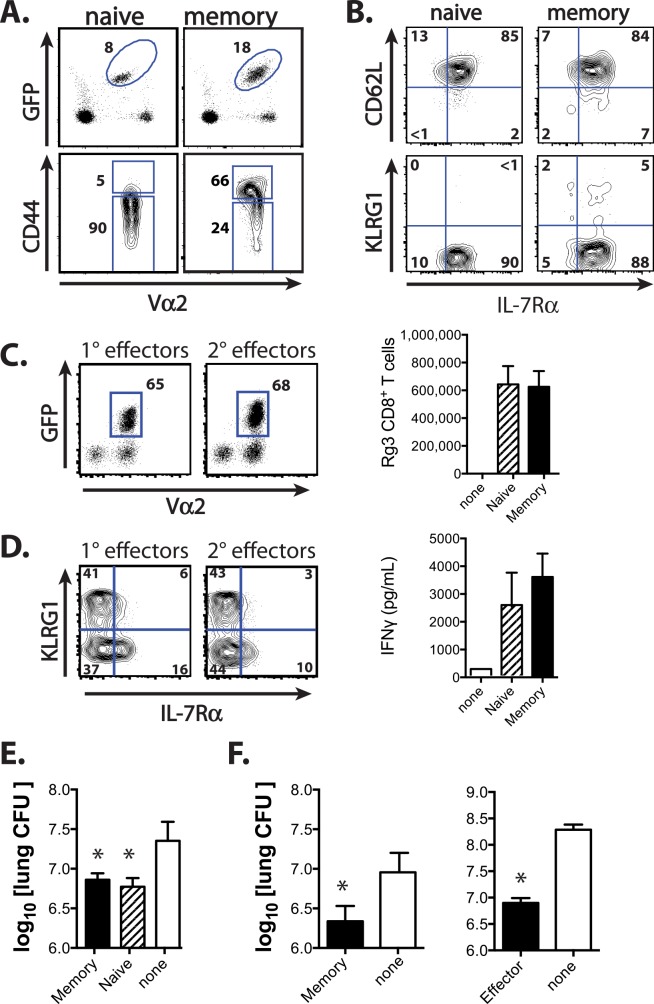
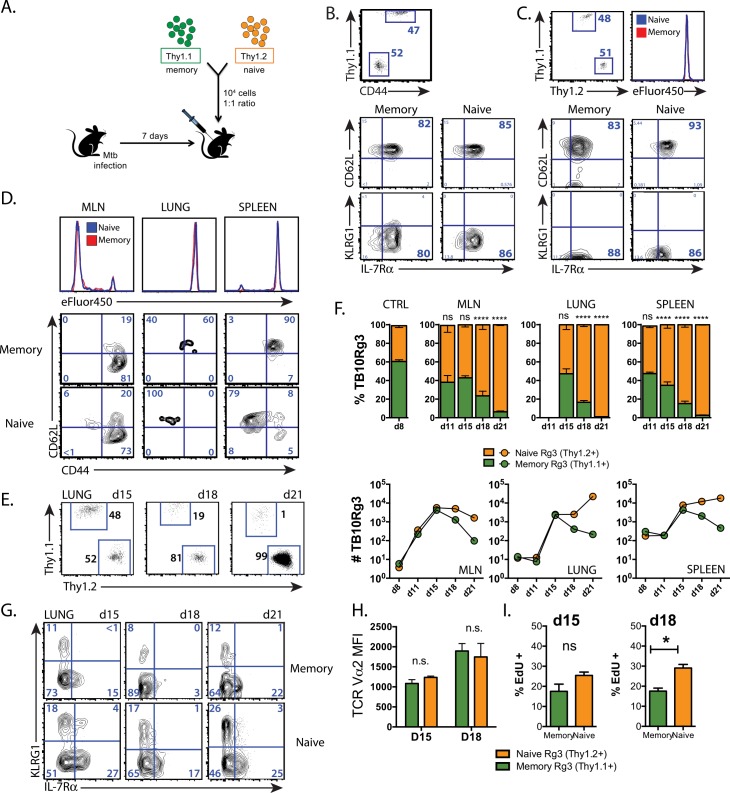
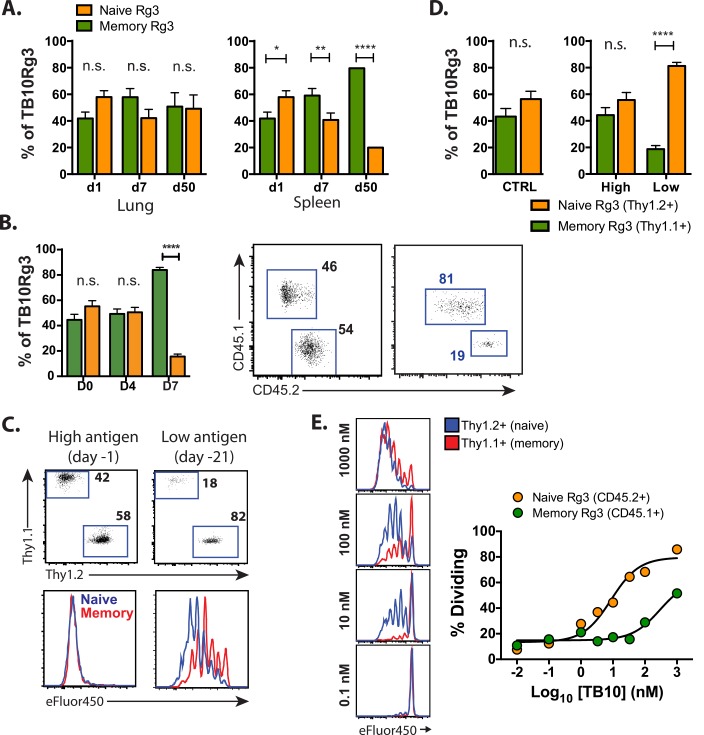
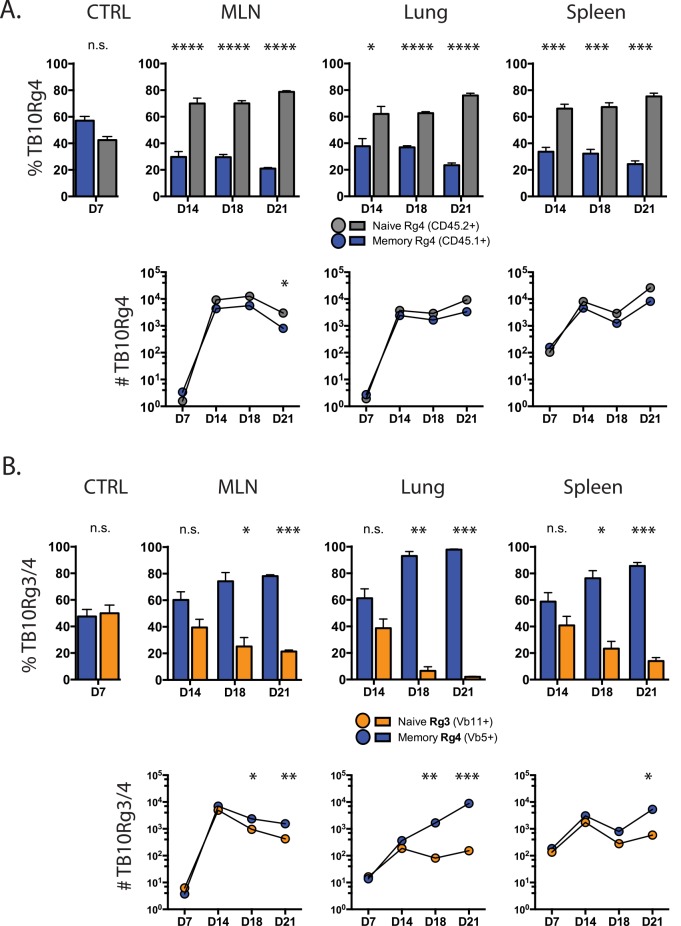
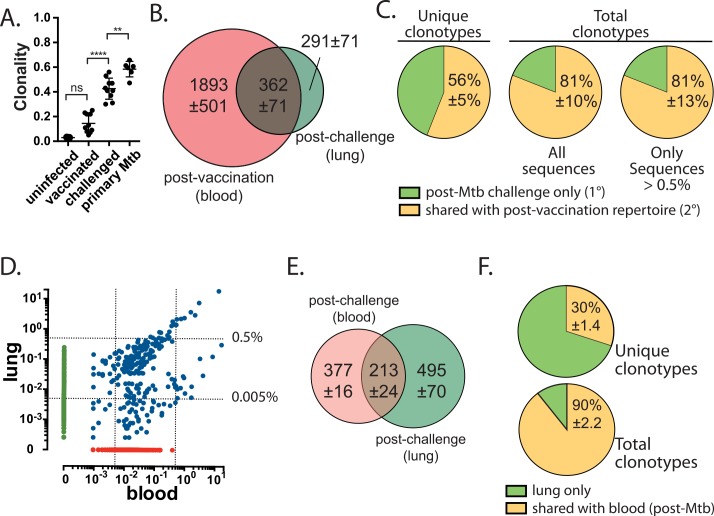
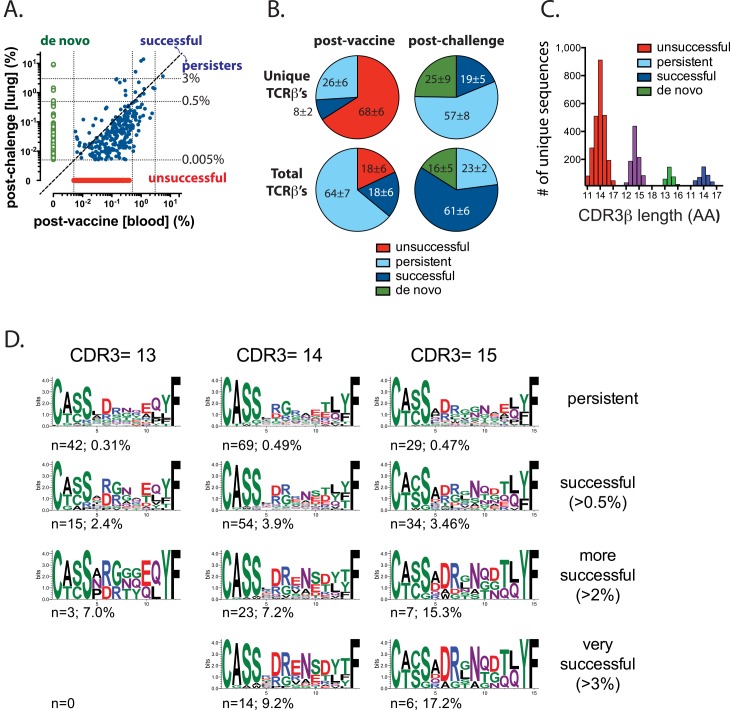
T cell vaccines against Mycobacterium tuberculosis (Mtb) and other pathogens are based on the principle that memory T cells rapidly generate effector responses upon challenge, leading to pathogen clearance. Despite eliciting a robust memory CD8+ T cell response to the immunodominant Mtb antigen TB10.4 (EsxH), we find the increased frequency of TB10.4-specific CD8+ T cells conferred by vaccination to be short-lived after Mtb challenge. To compare memory and naïve CD8+ T cell function during their response to Mtb, we track their expansions using TB10.4-specific retrogenic CD8+ T cells. We find that the primary (naïve) response outnumbers the secondary (memory) response during Mtb challenge, an effect moderated by increased TCR affinity. To determine whether the expansion of polyclonal memory T cells is restrained following Mtb challenge, we used TCRβ deep sequencing to track TB10.4-specific CD8+ T cells after vaccination and subsequent challenge in intact mice. Successful memory T cells, defined by their clonal expansion after Mtb challenge, express similar CDR3β sequences suggesting TCR selection by antigen. Thus, both TCR-dependent and -independent factors affect the fitness of memory CD8+ responses. The impaired expansion of the majority of memory T cell clonotypes may explain why some TB vaccines have not provided better protection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Swain SL, Weinberg AD, English M. CD4+ T cell subsets. Lymphokine secretion of memory cells and of effector cells that develop from precursors in vitro. J Immunol. 1990;144: 1788–1799. - PubMed
-
- Swain SL, Bradley LM, Croft M, Tonkonogy S, Atkins G, Weinberg AD, et al. Helper T-cell subsets: phenotype, function and the role of lymphokines in regulating their development. Immunol Rev. 1991;123: 115–144. - PubMed
-
- Ahmed R, Gray D. Immunological memory and protective immunity: understanding their relation. Science. 1996;272: 54–60. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
