

Mutations of Arabidopsis TBL32 and TBL33 Affect Xylan Acetylation and Secondary Wall Deposition
- PMID: 26745802
- PMCID: PMC4712945
- DOI: 10.1371/journal.pone.0146460
Mutations of Arabidopsis TBL32 and TBL33 Affect Xylan Acetylation and Secondary Wall Deposition
Abstract
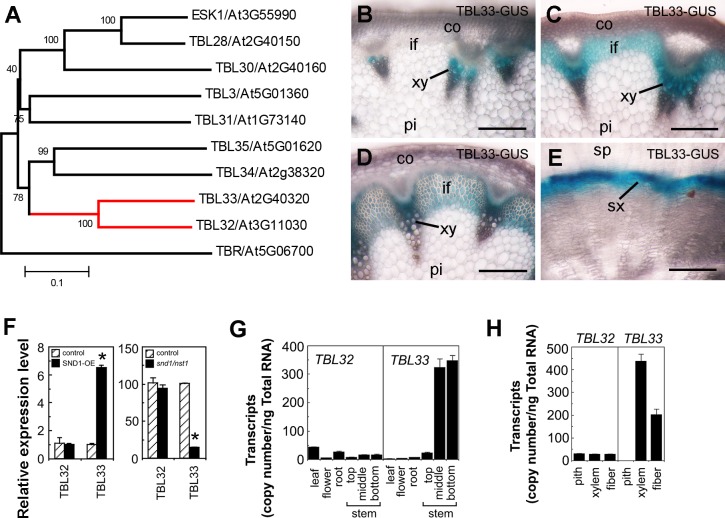
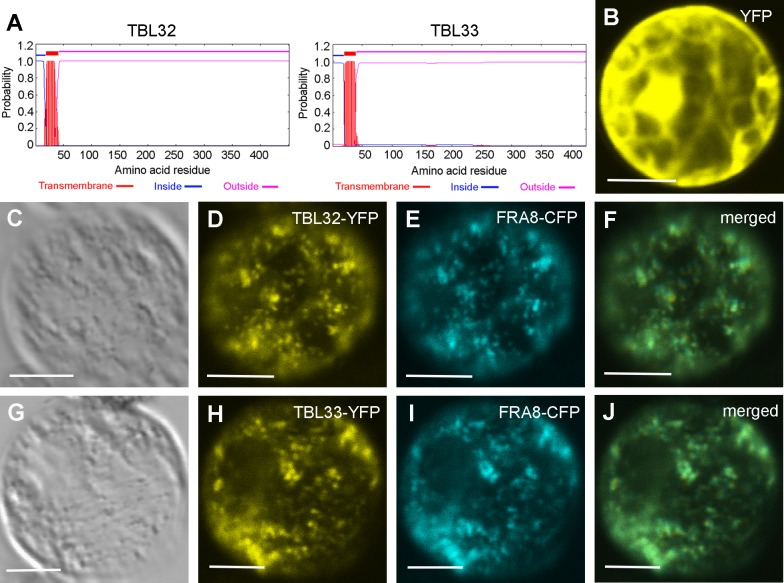
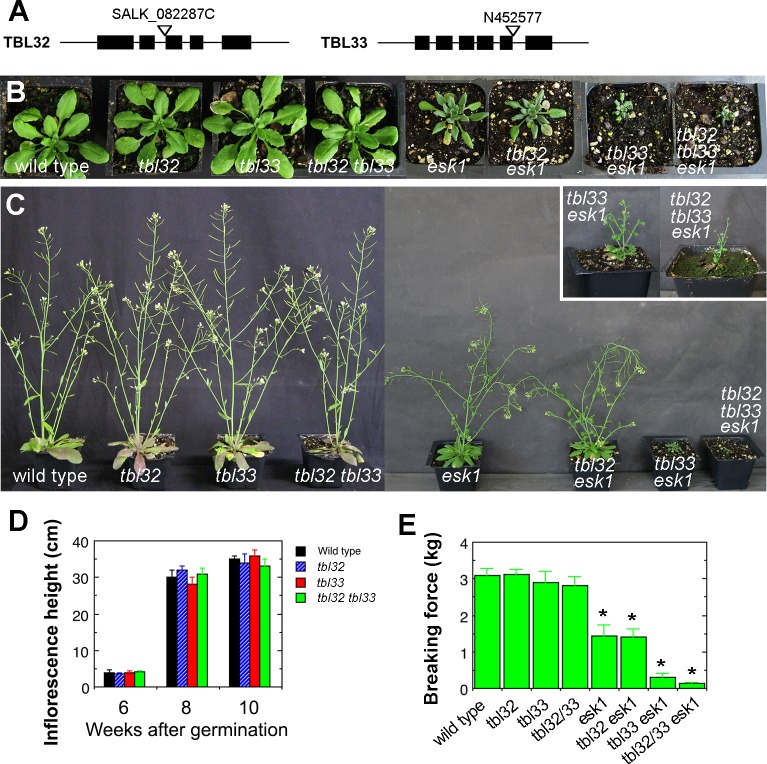
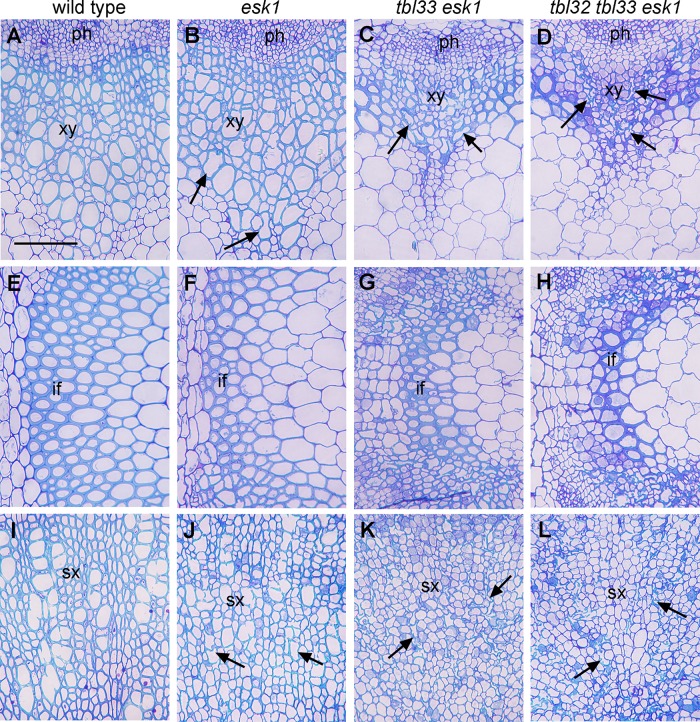
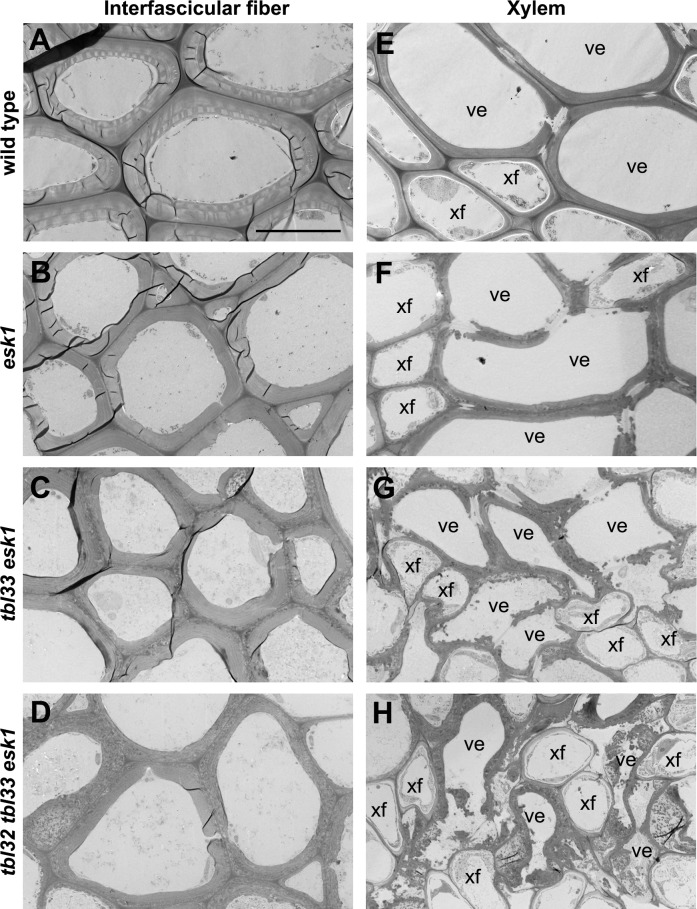
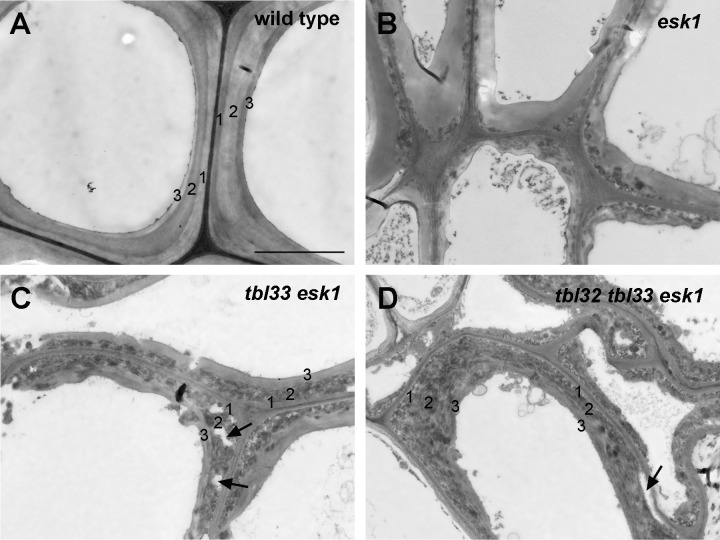
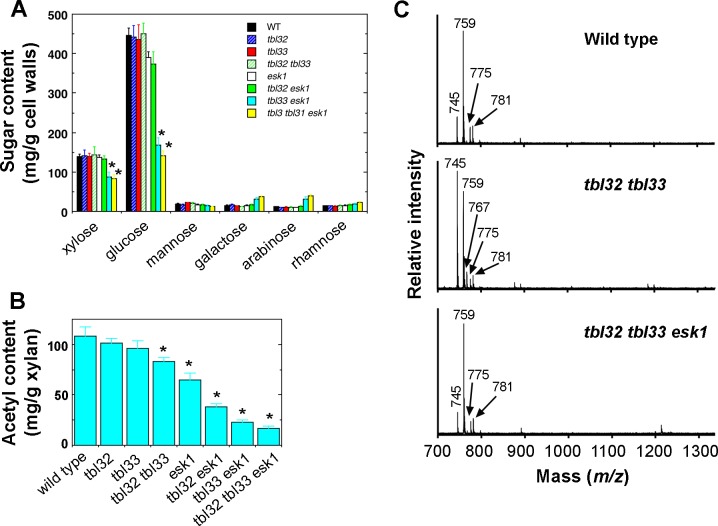
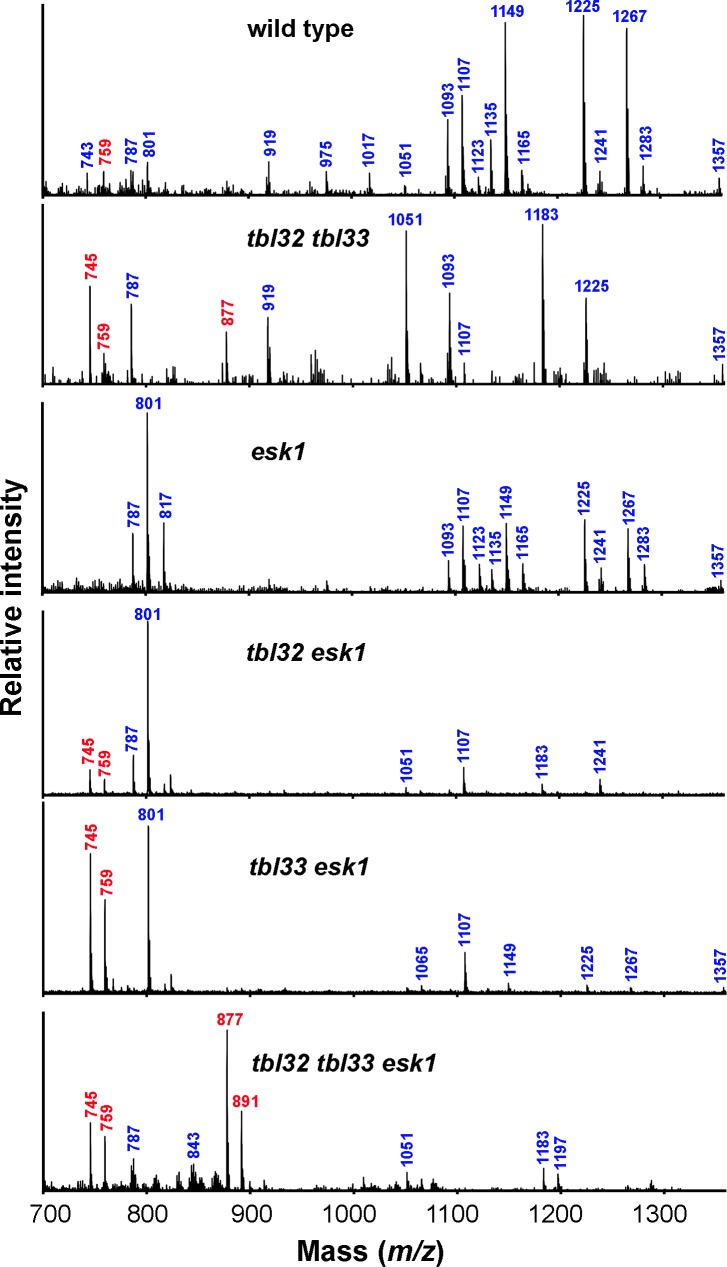
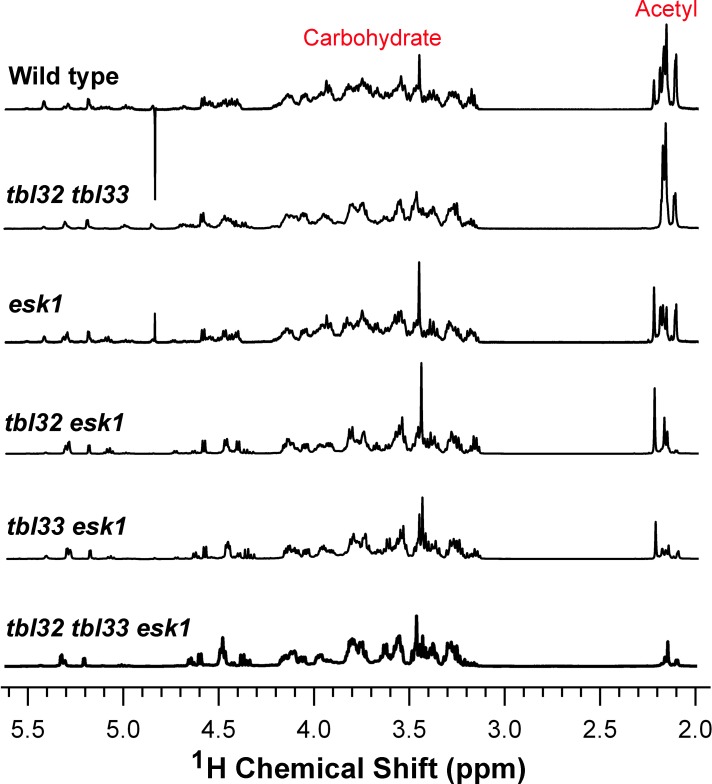
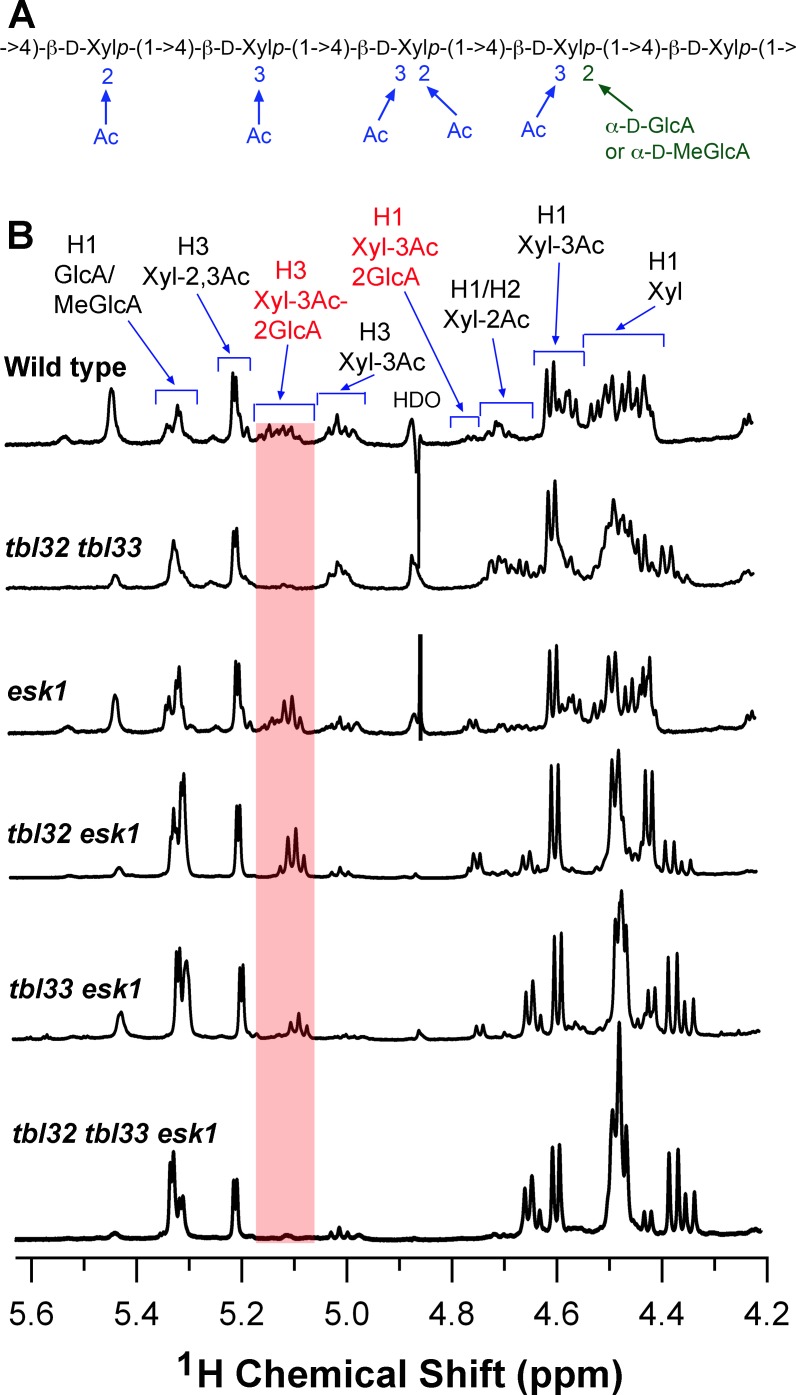
Xylan is a major acetylated polymer in plant lignocellulosic biomass and it can be mono- and di-acetylated at O-2 and O-3 as well as mono-acetylated at O-3 of xylosyl residues that is substituted with glucuronic acid (GlcA) at O-2. Based on the finding that ESK1, an Arabidopsis thaliana DUF231 protein, specifically mediates xylan 2-O- and 3-O-monoacetylation, we previously proposed that different acetyltransferase activities are required for regiospecific acetyl substitutions of xylan. Here, we demonstrate the functional roles of TBL32 and TBL33, two ESK1 close homologs, in acetyl substitutions of xylan. Simultaneous mutations of TBL32 and TBL33 resulted in a significant reduction in xylan acetyl content and endoxylanase digestion of the mutant xylan released GlcA-substituted xylooligomers without acetyl groups. Structural analysis of xylan revealed that the tbl32 tbl33 mutant had a nearly complete loss of 3-O-acetylated, 2-O-GlcA-substituted xylosyl residues. A reduction in 3-O-monoacetylated and 2,3-di-O-acetylated xylosyl residues was also observed. Simultaneous mutations of TBL32, TBL33 and ESK1 resulted in a severe reduction in xylan acetyl level down to 15% of that of the wild type, and concomitantly, severely collapsed vessels and stunted plant growth. In particular, the S2 layer of secondary walls in xylem vessels of tbl33 esk1 and tbl32 tbl33 esk1 exhibited an altered structure, indicating abnormal assembly of secondary wall polymers. These results demonstrate that TBL32 and TBL33 play an important role in xylan acetylation and normal deposition of secondary walls.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
