

Efficient expression of nuclear transgenes in the green alga Chlamydomonas: synthesis of an HIV antigen and development of a new selectable marker
- PMID: 26747175
- PMCID: PMC4766212
- DOI: 10.1007/s11103-015-0425-8
Efficient expression of nuclear transgenes in the green alga Chlamydomonas: synthesis of an HIV antigen and development of a new selectable marker
Abstract
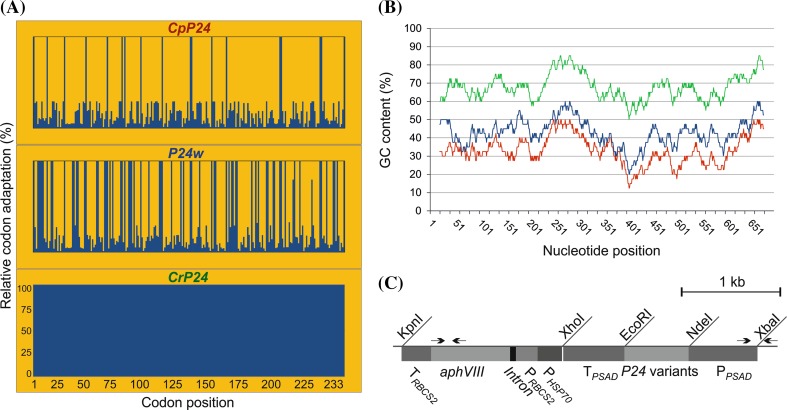
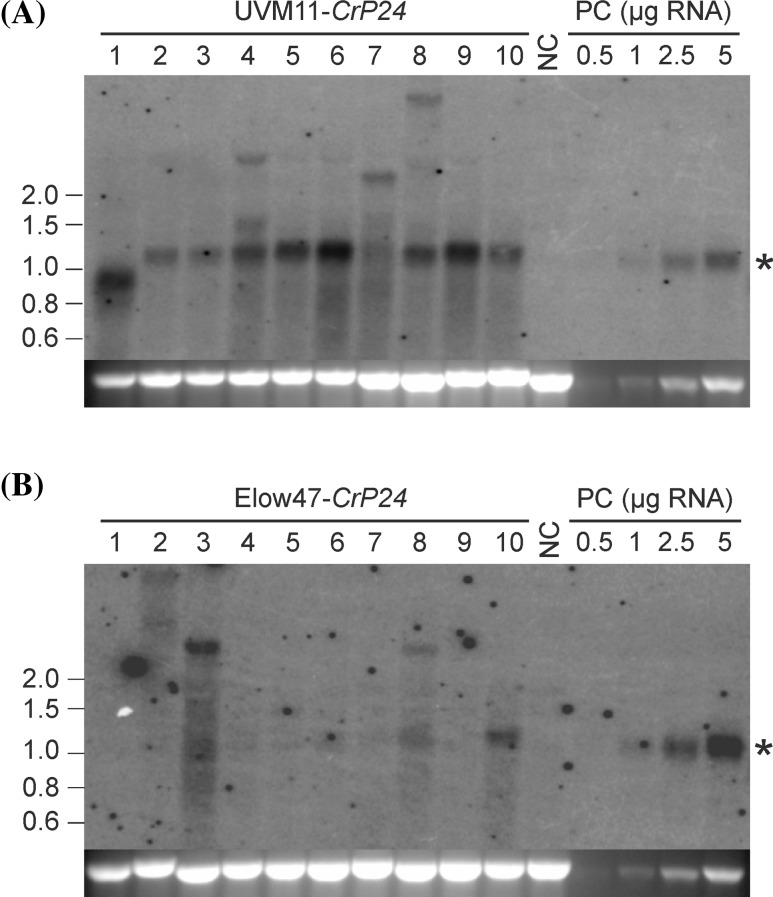
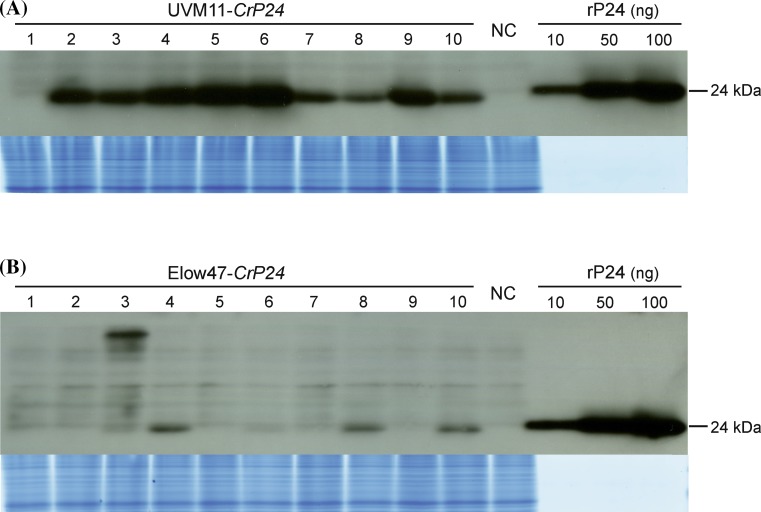
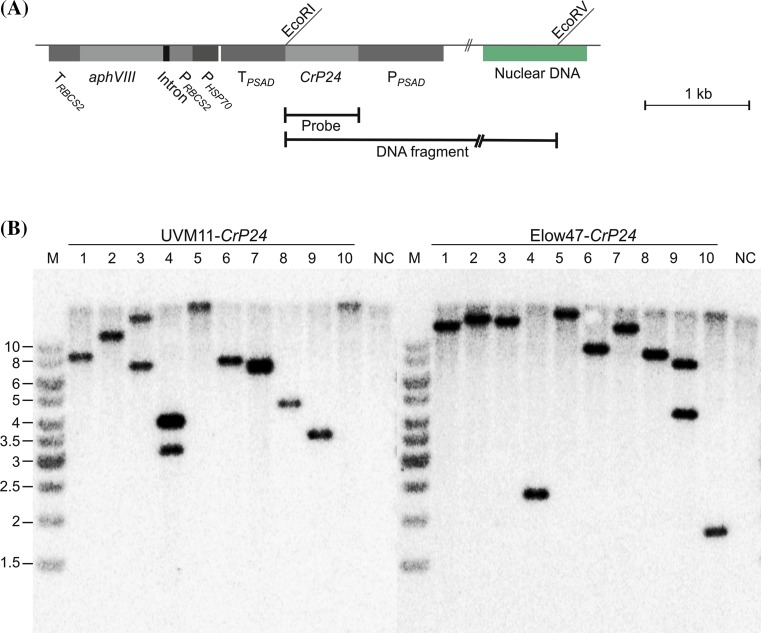
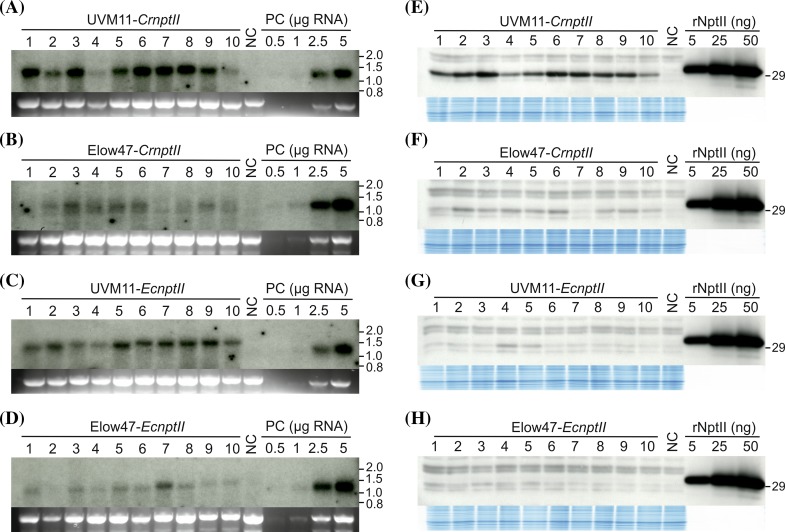
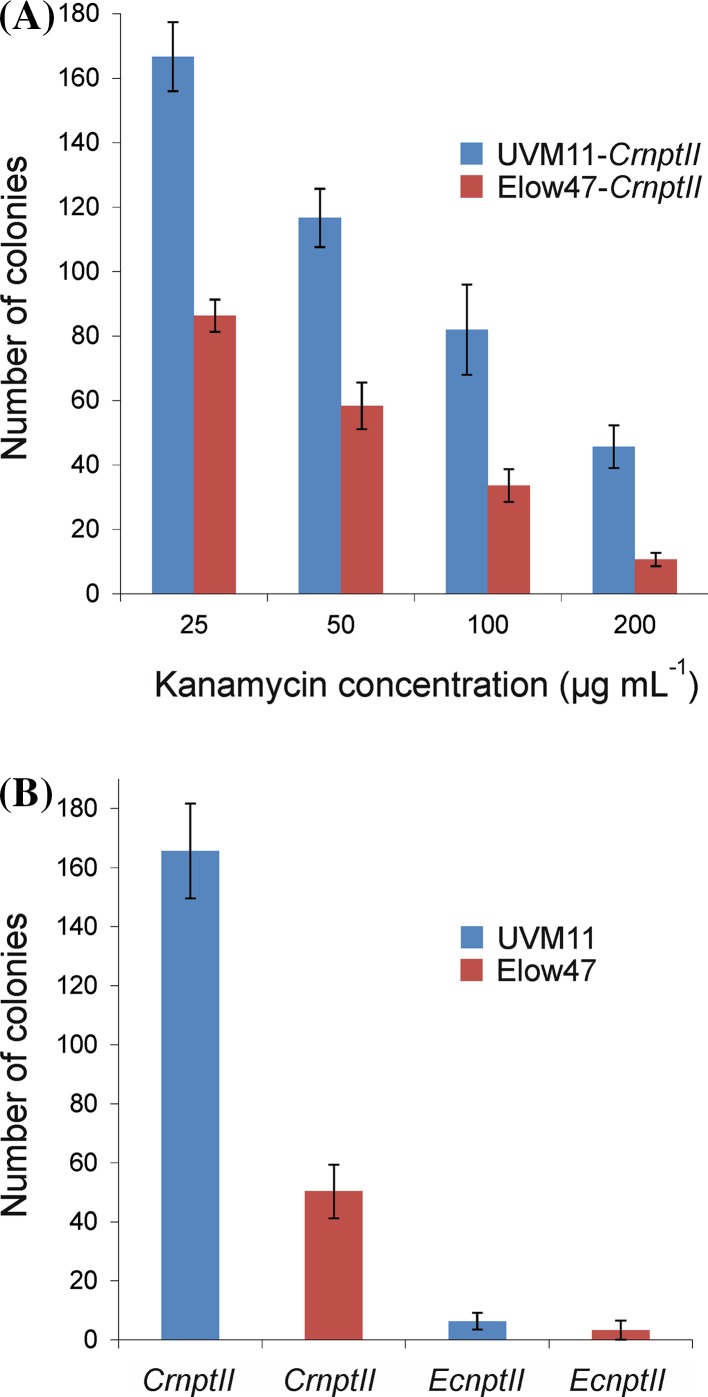
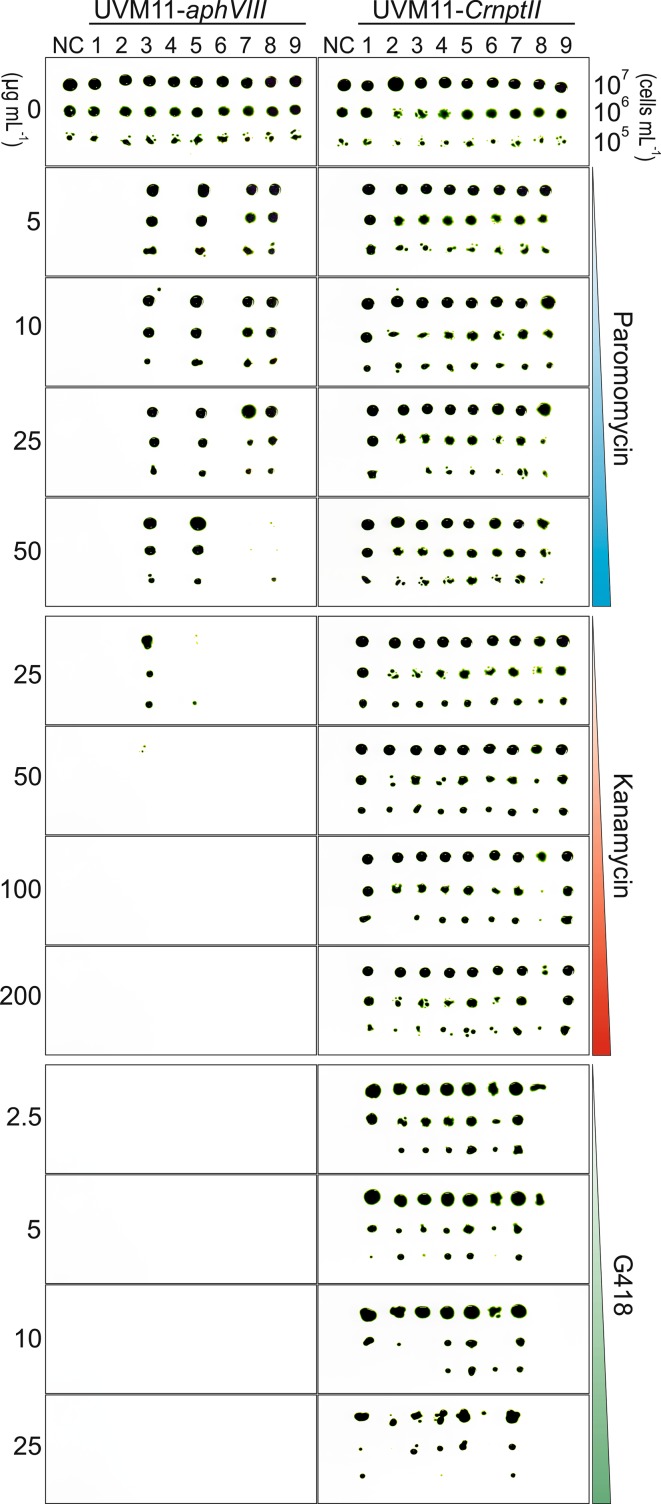
The unicellular green alga Chlamydomonas reinhardtii has become an invaluable model system in plant biology. There is also considerable interest in developing this microalga into an efficient production platform for biofuels, pharmaceuticals, green chemicals and industrial enzymes. However, the production of foreign proteins in the nucleocytosolic compartment of Chlamydomonas is greatly hampered by the inefficiency of transgene expression from the nuclear genome. We have recently addressed this limitation by isolating mutant algal strains that permit high-level transgene expression and by determining the contributions of GC content and codon usage to gene expression efficiency. Here we have applied these new tools and explored the potential of Chlamydomonas to produce a recombinant biopharmaceutical, the HIV antigen P24. We show that a codon-optimized P24 gene variant introduced into our algal expression strains give rise to recombinant protein accumulation levels of up to 0.25% of the total cellular protein. Moreover, in combination with an expression strain, a resynthesized nptII gene becomes a highly efficient selectable marker gene that facilitates the selection of transgenic algal clones at high frequency. By establishing simple principles of successful transgene expression, our data open up new possibilities for biotechnological research in Chlamydomonas.
Keywords: Antigen; Chlamydomonas reinhardtii; HIV; Molecular farming; Selectable marker gene; Transformation.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
