

Chloroplast Activity and 3'phosphadenosine 5'phosphate Signaling Regulate Programmed Cell Death in Arabidopsis
- PMID: 26747283
- PMCID: PMC4775142
- DOI: 10.1104/pp.15.01872
Chloroplast Activity and 3'phosphadenosine 5'phosphate Signaling Regulate Programmed Cell Death in Arabidopsis
Abstract
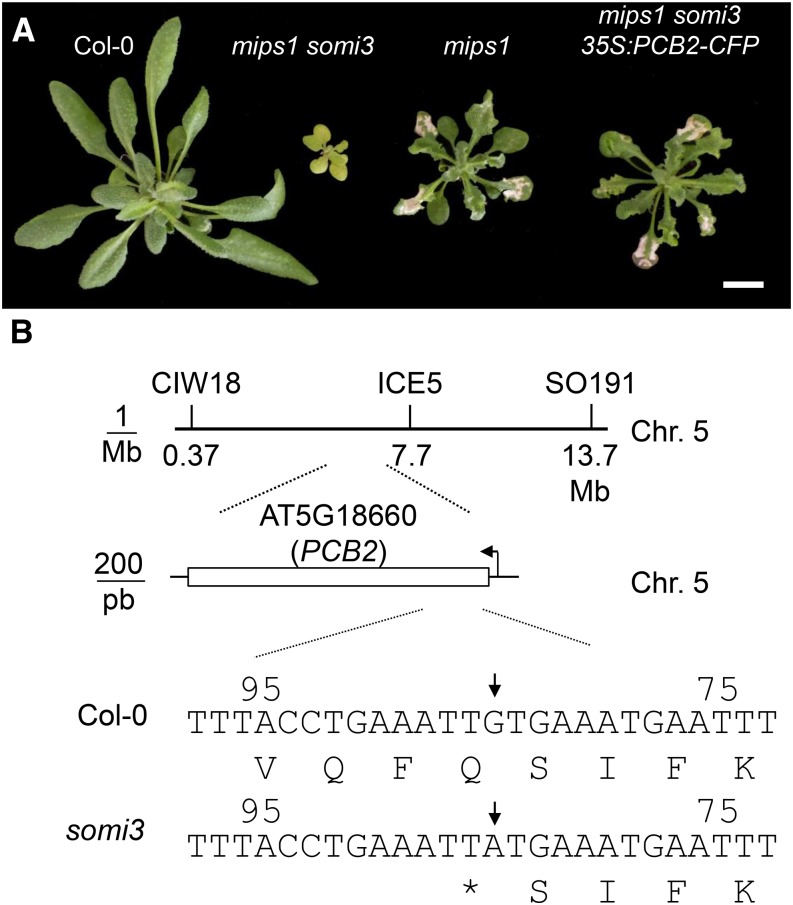
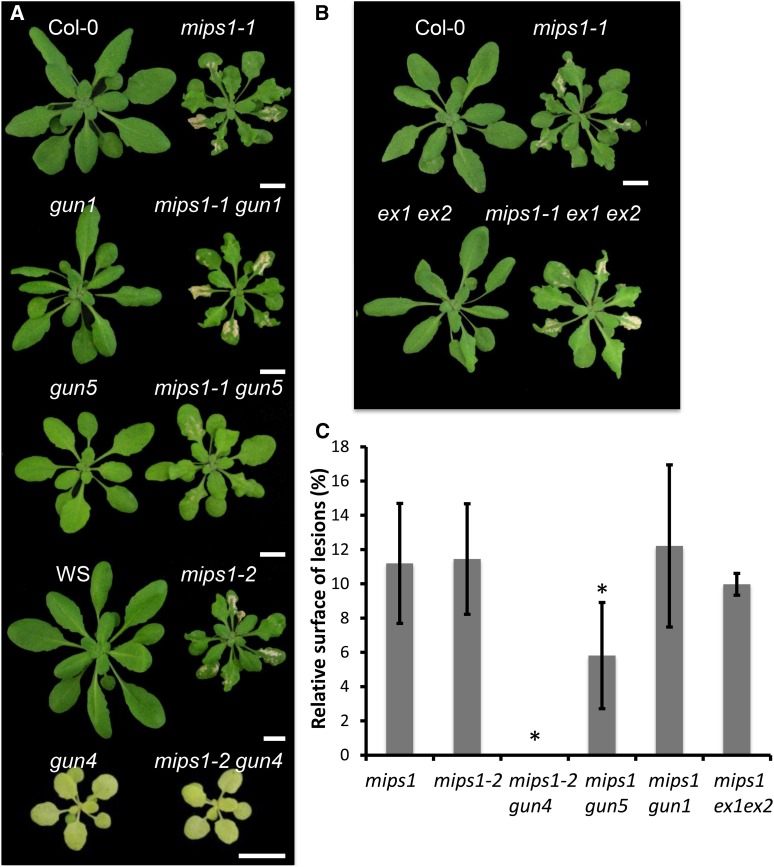
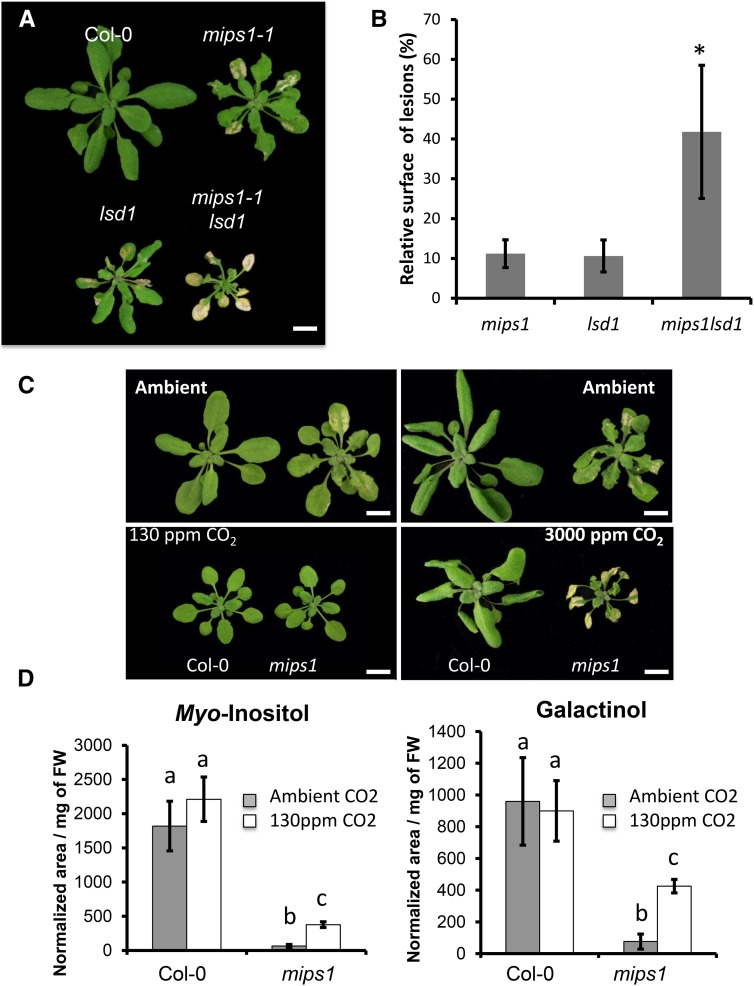
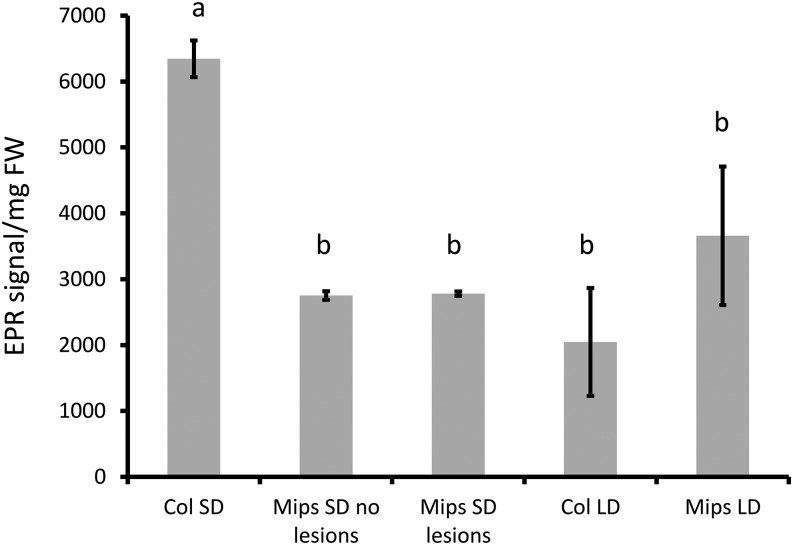
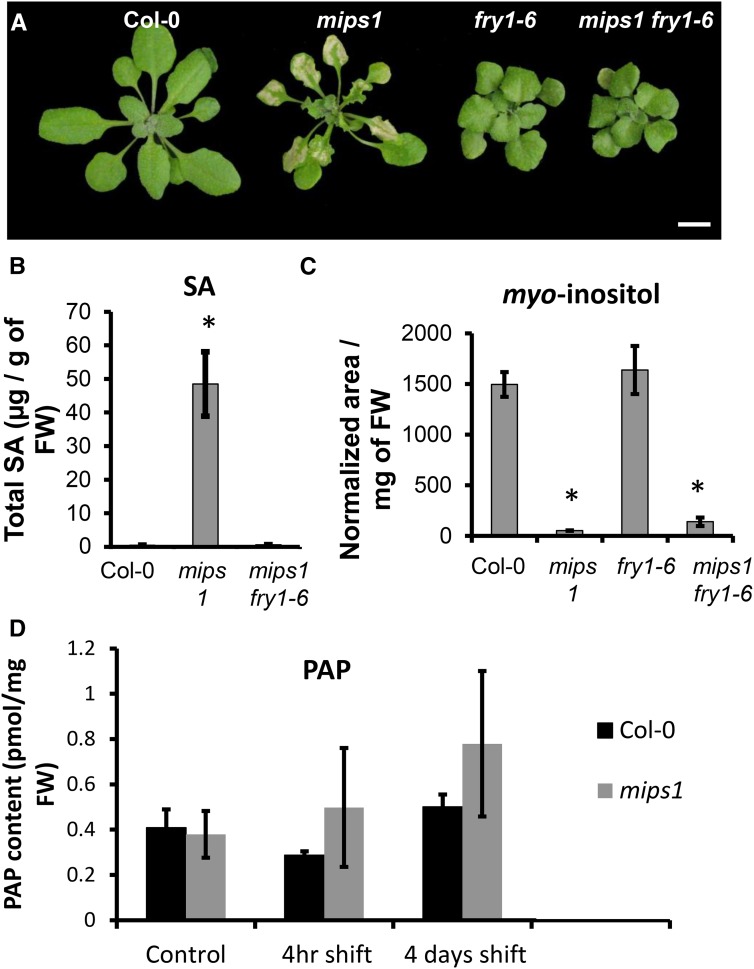
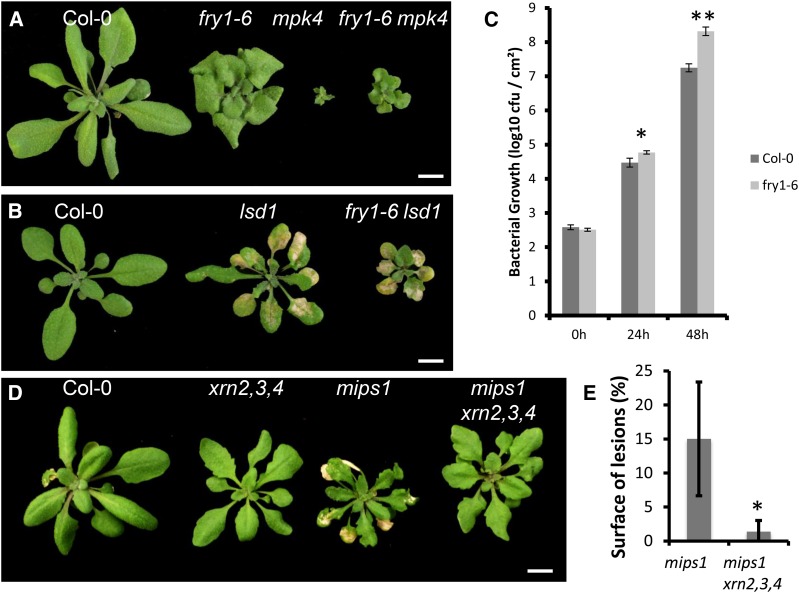
Programmed cell death (PCD) is a crucial process both for plant development and responses to biotic and abiotic stress. There is accumulating evidence that chloroplasts may play a central role during plant PCD as for mitochondria in animal cells, but it is still unclear whether they participate in PCD onset, execution, or both. To tackle this question, we have analyzed the contribution of chloroplast function to the cell death phenotype of the myoinositol phosphate synthase1 (mips1) mutant that forms spontaneous lesions in a light-dependent manner. We show that photosynthetically active chloroplasts are required for PCD to occur in mips1, but this process is independent of the redox state of the chloroplast. Systematic genetic analyses with retrograde signaling mutants reveal that 3'-phosphoadenosine 5'-phosphate, a chloroplast retrograde signal that modulates nuclear gene expression in response to stress, can inhibit cell death and compromises plant innate immunity via inhibition of the RNA-processing 5'-3' exoribonucleases. Our results provide evidence for the role of chloroplast-derived signal and RNA metabolism in the control of cell death and biotic stress response.
© 2016 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al. (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
-
- Aviv DH, Rustérucci C, Holt BF III, Dietrich RA, Parker JE, Dangl JL (2002) Runaway cell death, but not basal disease resistance, in lsd1 is SA- and NIM1/NPR1-dependent. Plant J 29: 381–391 - PubMed
-
- Bednarek P, Pislewska-Bednarek M, Svatos A, Schneider B, Doubsky J, Mansurova M, Humphry M, Consonni C, Panstruga R, Sanchez-Vallet A, et al. (2009) A glucosinolate metabolism pathway in living plant cells mediates broad-spectrum antifungal defense. Science 323: 101–106 - PubMed
-
- Bruggeman Q, Garmier M, de Bont L, Soubigou-Taconnat L, Mazubert C, Benhamed M, Raynaud C, Bergounioux C, Delarue M (2014) The polyadenylation factor subunit CLEAVAGE AND POLYADENYLATION SPECIFICITY FACTOR30: a key factor of programmed cell death and a regulator of immunity in Arabidopsis. Plant Physiol 165: 732–746 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
