

Deficiency of Starch Synthase IIIa and IVb Alters Starch Granule Morphology from Polyhedral to Spherical in Rice Endosperm
- PMID: 26747287
- PMCID: PMC4775109
- DOI: 10.1104/pp.15.01232
Deficiency of Starch Synthase IIIa and IVb Alters Starch Granule Morphology from Polyhedral to Spherical in Rice Endosperm
Abstract
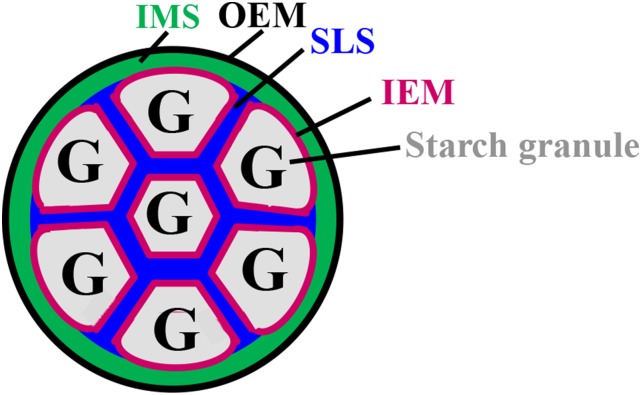
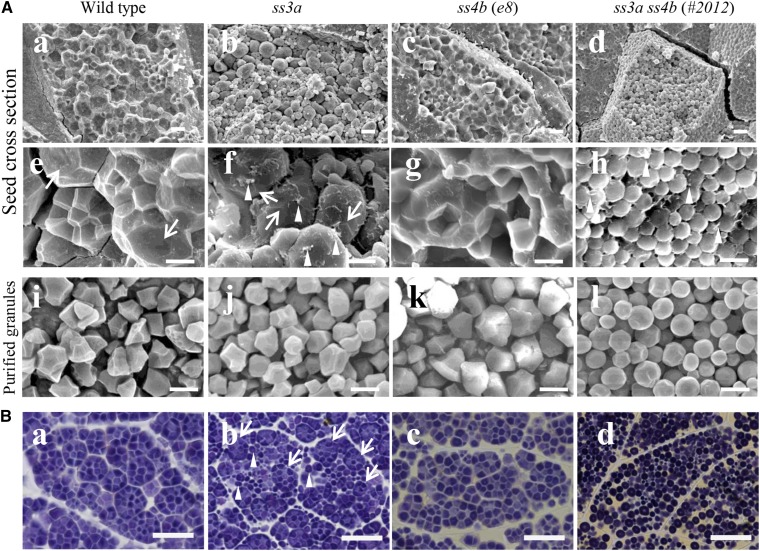
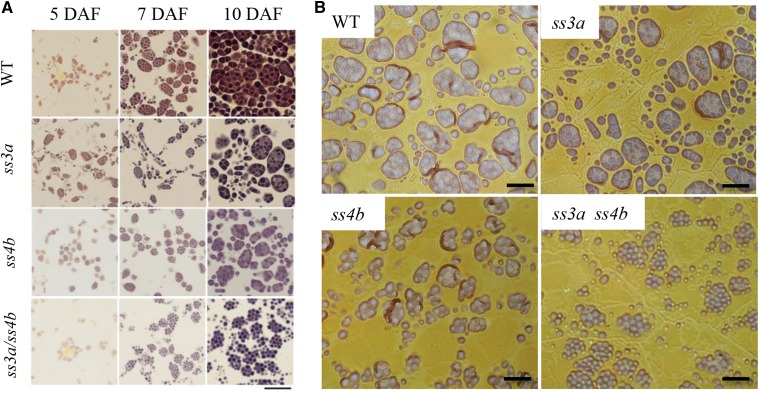
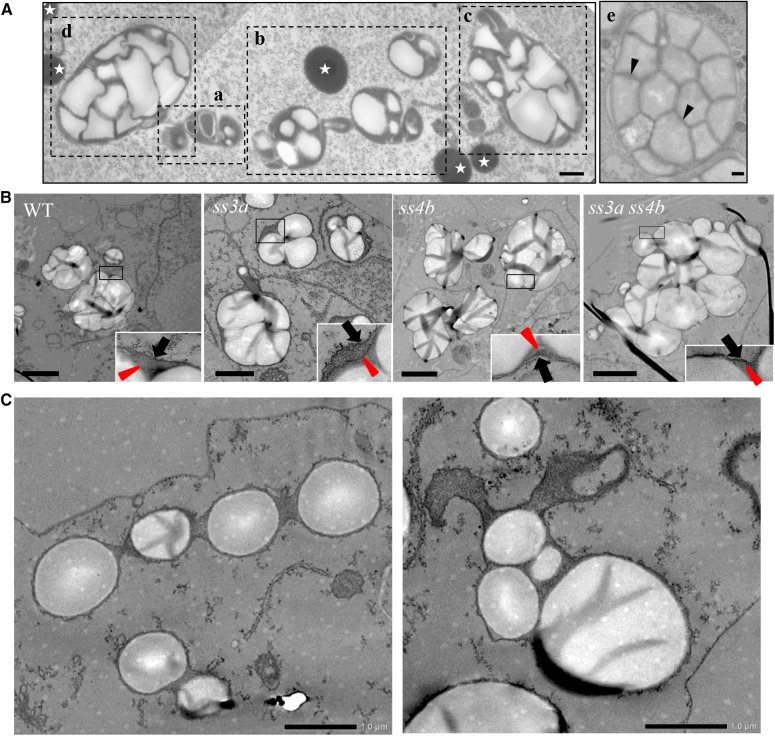
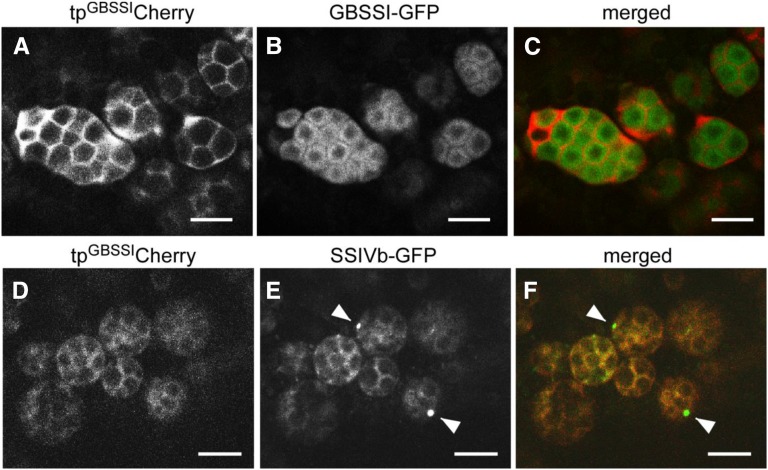
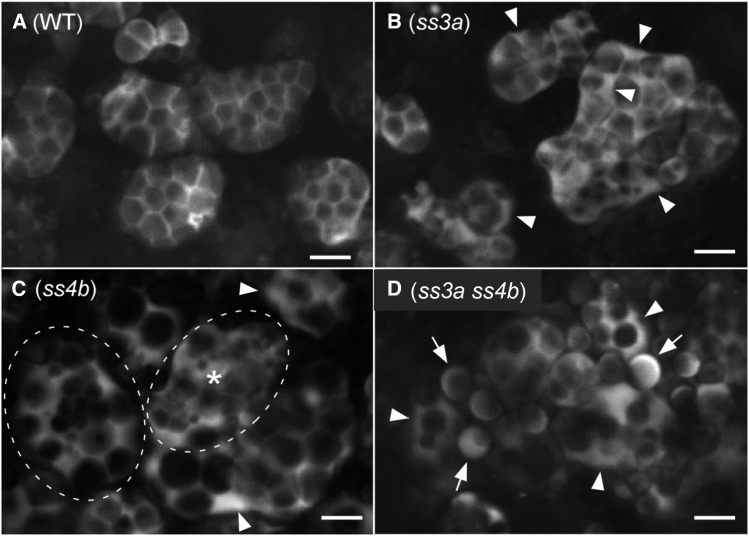
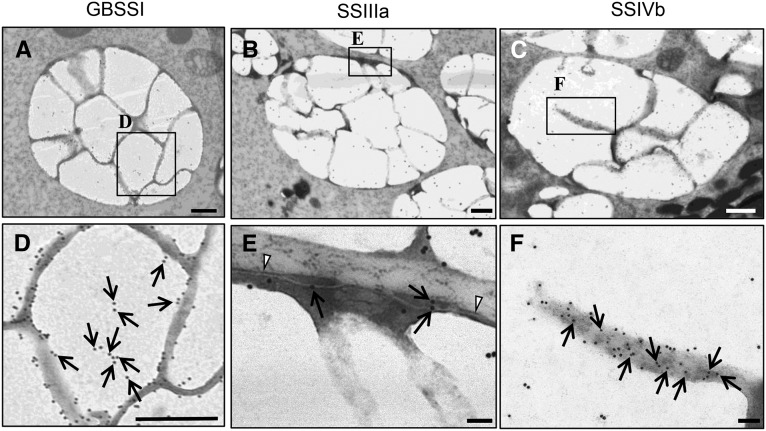
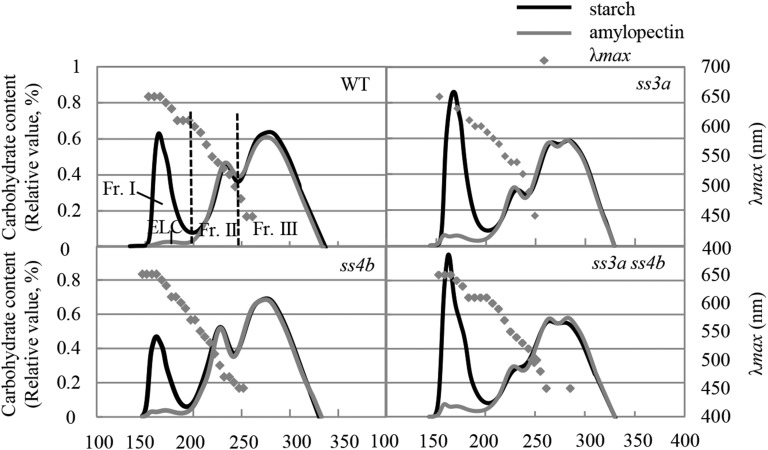
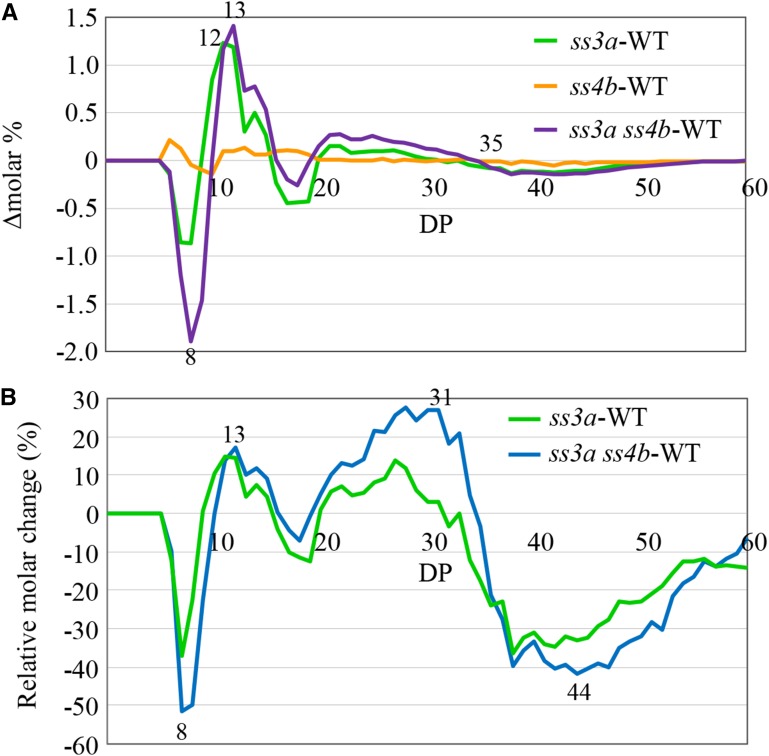
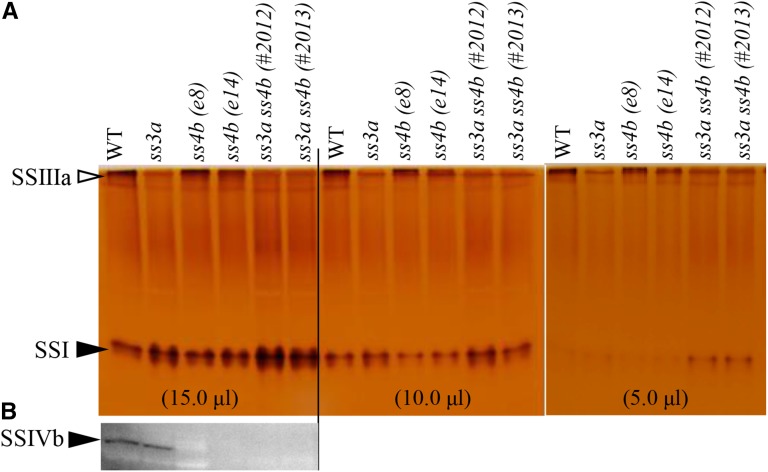
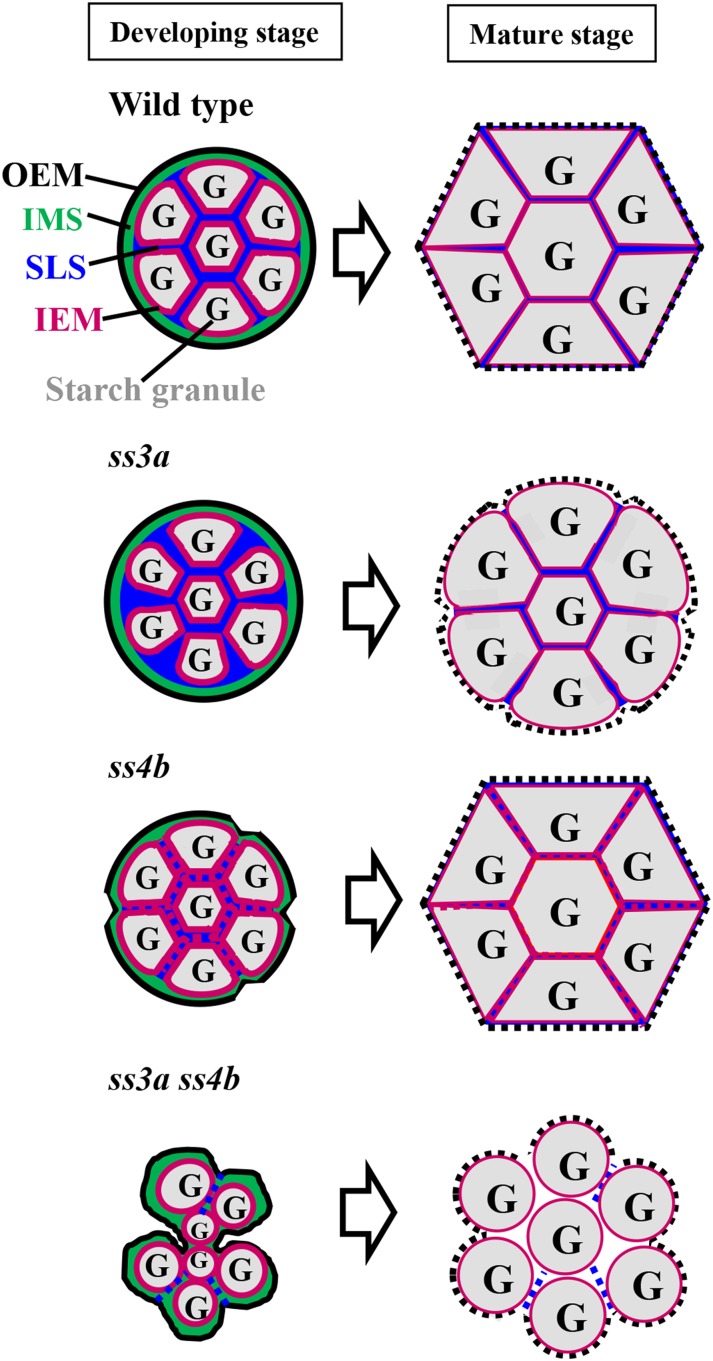
Starch granule morphology differs markedly among plant species. However, the mechanisms controlling starch granule morphology have not been elucidated. Rice (Oryza sativa) endosperm produces characteristic compound-type granules containing dozens of polyhedral starch granules within an amyloplast. Some other cereal species produce simple-type granules, in which only one starch granule is present per amyloplast. A double mutant rice deficient in the starch synthase (SS) genes SSIIIa and SSIVb (ss3a ss4b) produced spherical starch granules, whereas the parental single mutants produced polyhedral starch granules similar to the wild type. The ss3a ss4b amyloplasts contained compound-type starch granules during early developmental stages, and spherical granules were separated from each other during subsequent amyloplast development and seed dehydration. Analysis of glucan chain length distribution identified overlapping roles for SSIIIa and SSIVb in amylopectin chain synthesis, with a degree of polymerization of 42 or greater. Confocal fluorescence microscopy and immunoelectron microscopy of wild-type developing rice seeds revealed that the majority of SSIVb was localized between starch granules. Therefore, we propose that SSIIIa and SSIVb have crucial roles in determining starch granule morphology and in maintaining the amyloplast envelope structure. We present a model of spherical starch granule production.
© 2016 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Ball S, Colleoni C, Cenci U, Raj JN, Tirtiaux C (2011) The evolution of glycogen and starch metabolism in eukaryotes gives molecular clues to understand the establishment of plastid endosymbiosis. J Exp Bot 62: 1775–1801 - PubMed
-
- Delvallé D, Dumez S, Wattebled F, Roldán I, Planchot V, Berbezy P, Colonna P, Vyas D, Chatterjee M, Ball S, et al. (2005) Soluble starch synthase I: a major determinant for the synthesis of amylopectin in Arabidopsis thaliana leaves. Plant J 43: 398–412 - PubMed
-
- Deschamps P, Colleoni C, Nakamura Y, Suzuki E, Putaux JL, Buléon A, Haebel S, Ritte G, Steup M, Falcón LI, et al. (2008) Metabolic symbiosis and the birth of the plant kingdom. Mol Biol Evol 25: 536–548 - PubMed
-
- Fujita N, Hasegawa H, Taira T (2001) The isolation and characterization of a waxy mutant of diploid wheat (Triticum monococcum L.). Plant Sci 160: 595–602 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
