

Genomic Signatures of Selective Pressures and Introgression from Archaic Hominins at Human Innate Immunity Genes
- PMID: 26748513
- PMCID: PMC4716683
- DOI: 10.1016/j.ajhg.2015.11.014
Genomic Signatures of Selective Pressures and Introgression from Archaic Hominins at Human Innate Immunity Genes
Abstract
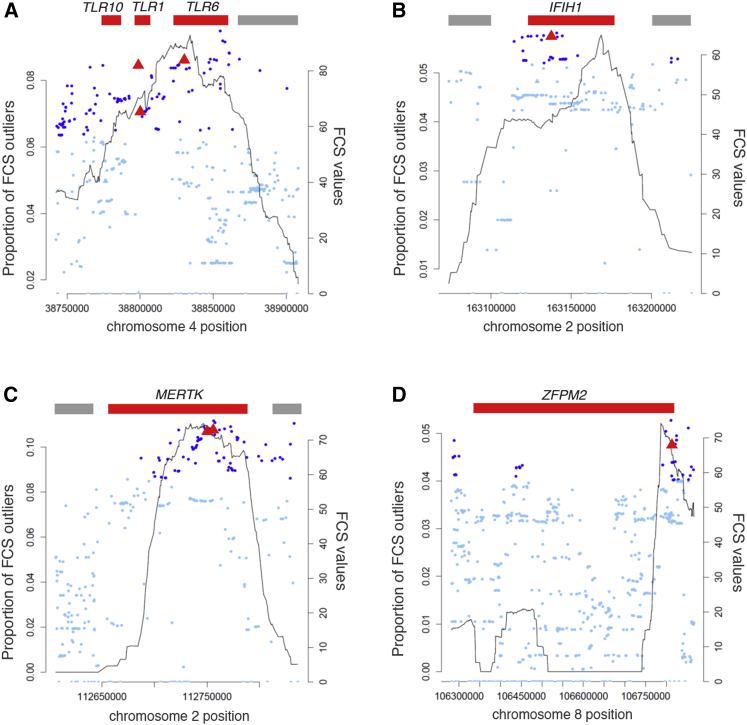
Human genes governing innate immunity provide a valuable tool for the study of the selective pressure imposed by microorganisms on host genomes. A comprehensive, genome-wide study of how selective constraints and adaptations have driven the evolution of innate immunity genes is missing. Using full-genome sequence variation from the 1000 Genomes Project, we first show that innate immunity genes have globally evolved under stronger purifying selection than the remainder of protein-coding genes. We identify a gene set under the strongest selective constraints, mutations in which are likely to predispose individuals to life-threatening disease, as illustrated by STAT1 and TRAF3. We then evaluate the occurrence of local adaptation and detect 57 high-scoring signals of positive selection at innate immunity genes, variation in which has been associated with susceptibility to common infectious or autoimmune diseases. Furthermore, we show that most adaptations targeting coding variation have occurred in the last 6,000-13,000 years, the period at which populations shifted from hunting and gathering to farming. Finally, we show that innate immunity genes present higher Neandertal introgression than the remainder of the coding genome. Notably, among the genes presenting the highest Neandertal ancestry, we find the TLR6-TLR1-TLR10 cluster, which also contains functional adaptive variation in Europeans. This study identifies highly constrained genes that fulfill essential, non-redundant functions in host survival and reveals others that are more permissive to change-containing variation acquired from archaic hominins or adaptive variants in specific populations-improving our understanding of the relative biological importance of innate immunity pathways in natural conditions.
Copyright © 2016 The American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Figures



Comment in
-
TLRs of Our Fathers.Immunity. 2016 Feb 16;44(2):218-20. doi: 10.1016/j.immuni.2016.02.003. Immunity. 2016. PMID: 26885854
References
-
- Casanova J.L., Abel L., Quintana-Murci L. Immunology taught by human genetics. Cold Spring Harb. Symp. Quant. Biol. 2013;78:157–172. - PubMed
-
- Chapman S.J., Hill A.V. Human genetic susceptibility to infectious disease. Nat. Rev. Genet. 2012;13:175–188. - PubMed
-
- Barreiro L.B., Quintana-Murci L. From evolutionary genetics to human immunology: how selection shapes host defence genes. Nat. Rev. Genet. 2010;11:17–30. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
