

MDM2 Associates with Polycomb Repressor Complex 2 and Enhances Stemness-Promoting Chromatin Modifications Independent of p53
- PMID: 26748827
- PMCID: PMC6284523
- DOI: 10.1016/j.molcel.2015.12.008
MDM2 Associates with Polycomb Repressor Complex 2 and Enhances Stemness-Promoting Chromatin Modifications Independent of p53
Abstract
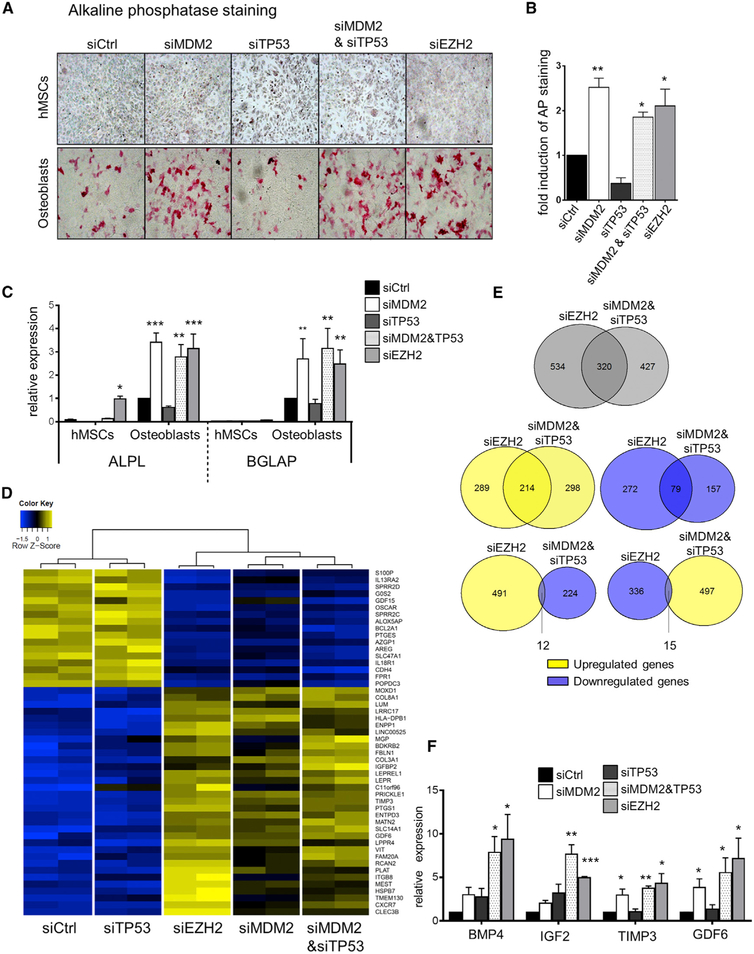
The MDM2 oncoprotein ubiquitinates and antagonizes p53 but may also carry out p53-independent functions. Here we report that MDM2 is required for the efficient generation of induced pluripotent stem cells (iPSCs) from murine embryonic fibroblasts, in the absence of p53. Similarly, MDM2 depletion in the context of p53 deficiency also promoted the differentiation of human mesenchymal stem cells and diminished clonogenic survival of cancer cells. Most of the MDM2-controlled genes also responded to the inactivation of the Polycomb Repressor Complex 2 (PRC2) and its catalytic component EZH2. MDM2 physically associated with EZH2 on chromatin, enhancing the trimethylation of histone 3 at lysine 27 and the ubiquitination of histone 2A at lysine 119 (H2AK119) at its target genes. Removing MDM2 simultaneously with the H2AK119 E3 ligase Ring1B/RNF2 further induced these genes and synthetically arrested cell proliferation. In conclusion, MDM2 supports the Polycomb-mediated repression of lineage-specific genes, independent of p53.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures






Comment in
-
Outside the p53 RING: Transcription Regulation by Chromatin-Bound MDM2.Mol Cell. 2016 Jun 16;62(6):805-807. doi: 10.1016/j.molcel.2016.05.035. Mol Cell. 2016. PMID: 27315554
References
-
- Armstrong JF, Kaufman MH, Harrison DJ, and Clarke AR (1995). High-frequency developmental abnormalities in p53-deficient mice. Curr. Biol. 5, 931–936. - PubMed
-
- Ben-Saadon R, Zaaroor D, Ziv T, and Ciechanover A (2006). The polycomb protein Ring1B generates self atypical mixed ubiquitin chains required for its in vitro histone H2A ligase activity. Mol. Cell 24, 701–711. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
