

The atypical IκB protein IκB(NS) is important for Toll-like receptor-induced interleukin-10 production in B cells
- PMID: 26749055
- PMCID: PMC4799890
- DOI: 10.1111/imm.12578
The atypical IκB protein IκB(NS) is important for Toll-like receptor-induced interleukin-10 production in B cells
Abstract
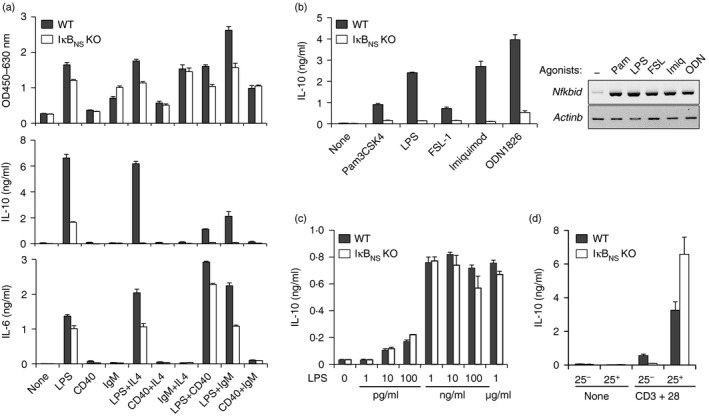
Although a major function of B cells is to mediate humoral immunity by producing antigen-specific antibodies, a specific subset of B cells is important for immune suppression, which is mainly mediated by the secretion of the anti-inflammatory cytokine interleukin-10 (IL-10). However, the mechanism by which IL-10 is induced in B cells has not been fully elucidated. Here, we report that IκBNS , an inducible nuclear IκB protein, is important for Toll-like receptor (TLR)-mediated IL-10 production in B cells. Studies using IκB(NS) knockout mice revealed that the number of IL-10-producing B cells is reduced in IκB(NS)(-/-) spleens and that the TLR-mediated induction of cytoplasmic IL-10-positive cells and IL-10 secretion in B cells are impaired in the absence of IκB(NS). The impairment of IL-10 production by a lack of IκB(NS) was not observed in TLR-triggered macrophages or T-cell-receptor-stimulated CD4(+) CD25(+) T cells. In addition, IκB(NS)-deficient B cells showed reduced expression of Prdm1 and Irf4 and failed to generate IL-10(+) CD138(+) plasmablasts. These results suggest that IκB(NS) is selectively required for IL-10 production in B cells responding to TLR signals, so defining an additional role for IκB(NS) in the control of the B-cell-mediated immune responses.
Keywords: IκBNS; Toll-like receptors; interleukin-10-producing B cells.
© 2016 John Wiley & Sons Ltd.
Figures





References
-
- Booth J, Wilson H, Jimbo S, Mutwiri G. Modulation of B cell responses by Toll‐like receptors. Cell Tissue Res 2011; 343:131–40. - PubMed
-
- Bernasconi NL, Onai N, Lanzavecchia A. A role for Toll‐like receptors in acquired immunity: up‐regulation of TLR9 by BCR triggering in naive B cells and constitutive expression in memory B cells. Blood 2003; 101:4500–4. - PubMed
-
- Ruprecht CR, Lanzavecchia A. Toll‐like receptor stimulation as a third signal required for activation of human naive B cells. Eur J Immunol 2006; 36:810–16. - PubMed
-
- Peng SL. Signaling in B cells via Toll‐like receptors. Curr Opin Immunol 2005; 17:230–6. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
