

Moderate (2%, v/v) Ethanol Feeding Alters Hepatic Wound Healing after Acute Carbon Tetrachloride Exposure in Mice
- PMID: 26751492
- PMCID: PMC4808799
- DOI: 10.3390/biom6010005
Moderate (2%, v/v) Ethanol Feeding Alters Hepatic Wound Healing after Acute Carbon Tetrachloride Exposure in Mice
Abstract
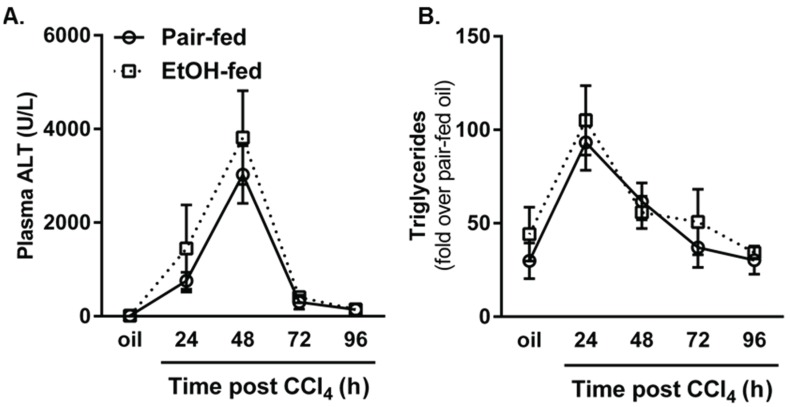
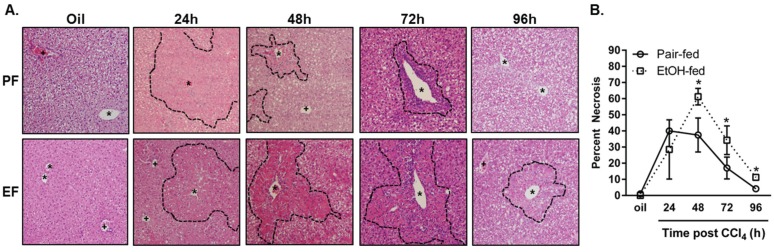
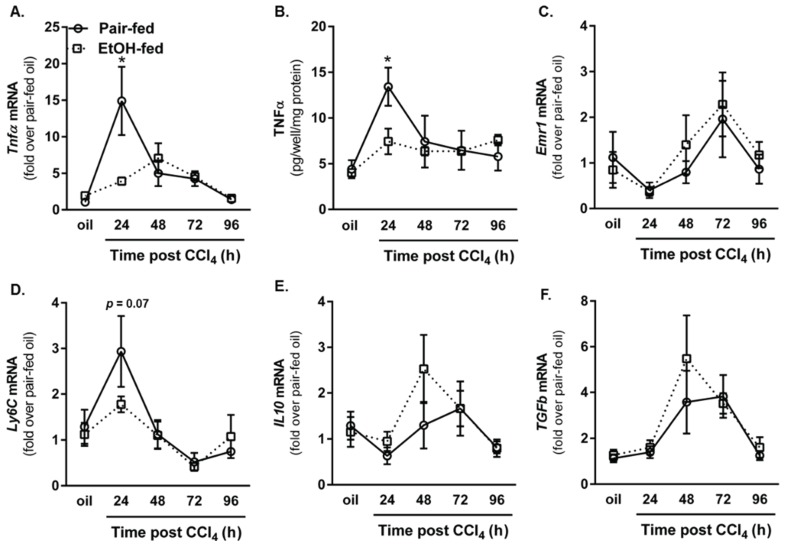
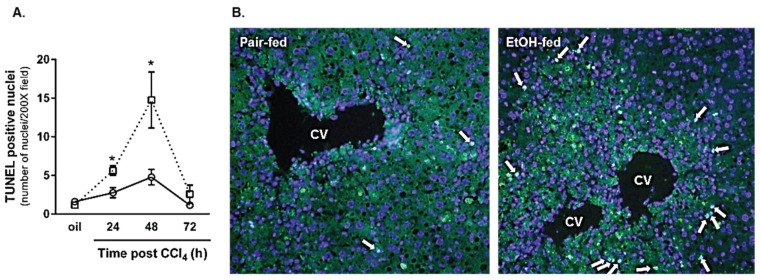
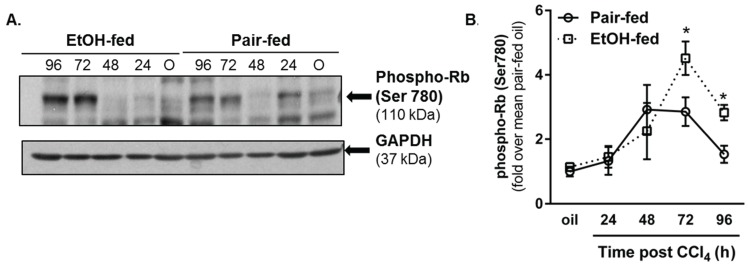
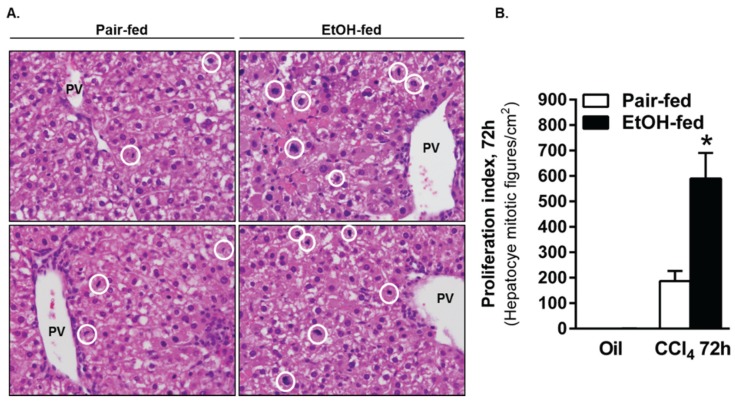
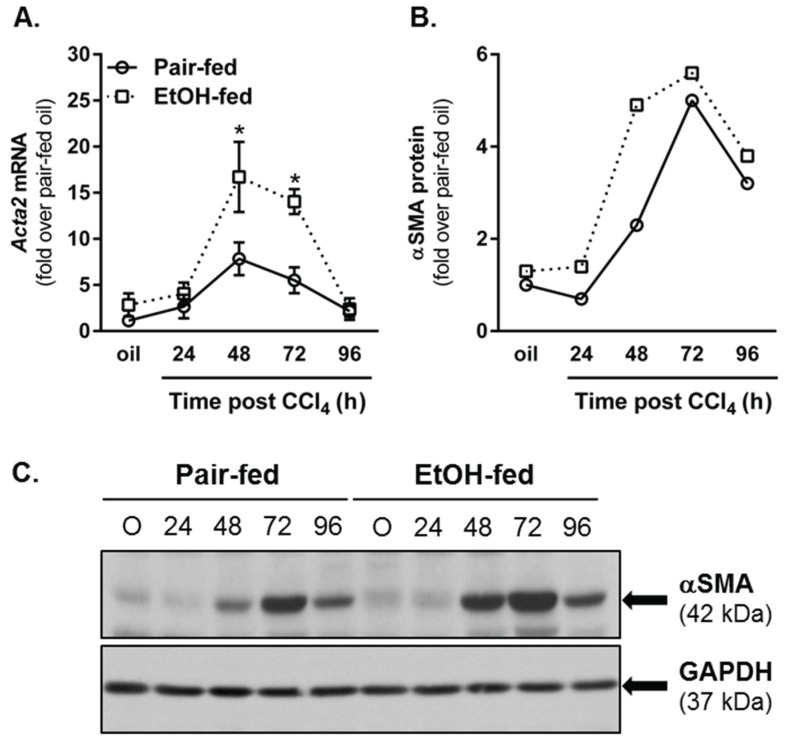
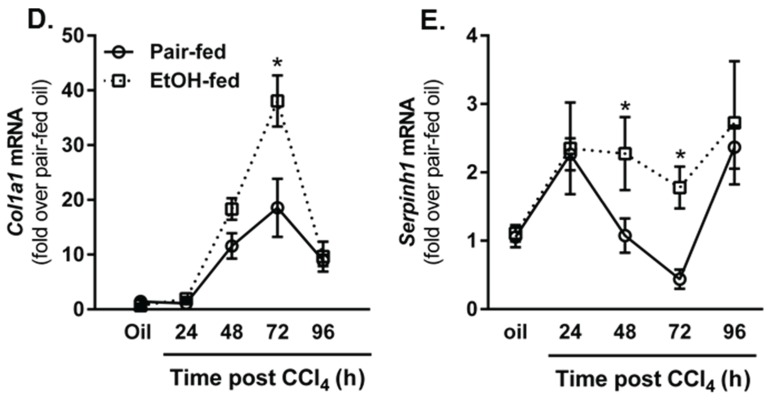
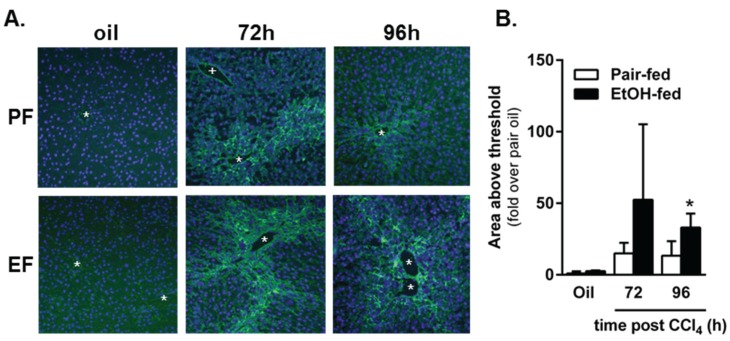
Wound healing consists of three overlapping phases: inflammation, proliferation, and matrix synthesis and remodeling. Prolonged alcohol abuse can cause liver fibrosis due to deregulated matrix remodeling. Previous studies demonstrated that moderate ethanol feeding enhances liver fibrogenic markers and frank fibrosis independent of differences in CCl₄-induced liver injury. Our objective was to determine whether or not other phases of the hepatic wound healing response were affected by moderate ethanol after CCl₄ exposure. Mice were fed moderate ethanol (2% v/v) for two days and then were exposed to CCl₄ and euthanized 24-96 h later. Liver injury was not different between pair- and ethanol-fed mice; however, removal of necrotic tissue was delayed after CCl₄-induced liver injury in ethanol-fed mice. Inflammation, measured by TNFα mRNA and protein and hepatic Ly6c transcript accumulation, was reduced and associated with enhanced hepatocyte apoptosis after ethanol feeding. Hepatocytes entered the cell cycle equivalently in pair- and ethanol-fed mice after CCl₄ exposure, but hepatocyte proliferation was prolonged in livers from ethanol-fed mice. CCl₄-induced hepatic stellate cell activation was increased and matrix remodeling was prolonged in ethanol-fed mice compared to controls. Taken together, moderate ethanol affected each phase of the wound healing response to CCl₄. These data highlight previously unknown effects of moderate ethanol exposure on hepatic wound healing after acute hepatotoxicant exposure.
Keywords: carbon tetrachloride; ethanol; fibrosis; inflammation; liver regeneration; matrix remodeling; wound healing.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
