

Modulation of the human gut microbiota by dietary fibres occurs at the species level
- PMID: 26754945
- PMCID: PMC4709873
- DOI: 10.1186/s12915-015-0224-3
Modulation of the human gut microbiota by dietary fibres occurs at the species level
Abstract
Background: Dietary intake of specific non-digestible carbohydrates (including prebiotics) is increasingly seen as a highly effective approach for manipulating the composition and activities of the human gut microbiota to benefit health. Nevertheless, surprisingly little is known about the global response of the microbial community to particular carbohydrates. Recent in vivo dietary studies have demonstrated that the species composition of the human faecal microbiota is influenced by dietary intake. There is now potential to gain insights into the mechanisms involved by using in vitro systems that produce highly controlled conditions of pH and substrate supply.
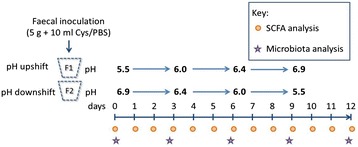
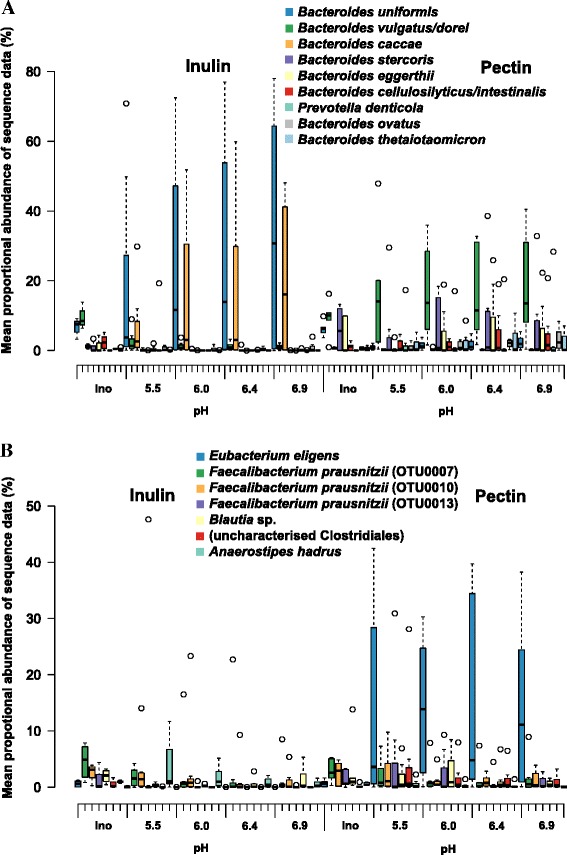
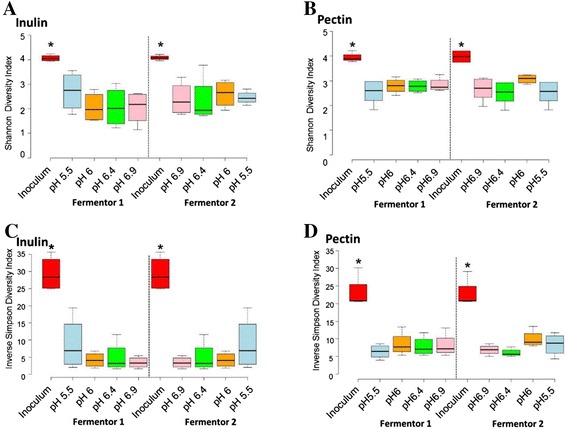
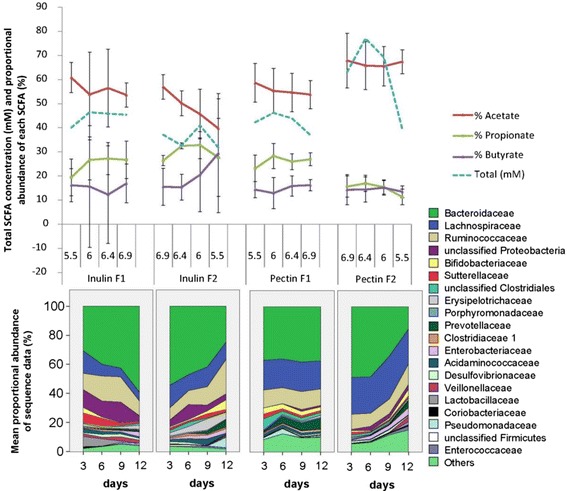
Results: We supplied two alternative non-digestible polysaccharides as energy sources to three different human gut microbial communities in anaerobic, pH-controlled continuous-flow fermentors. Community analysis showed that supply of apple pectin or inulin resulted in the highly specific enrichment of particular bacterial operational taxonomic units (OTUs; based on 16S rRNA gene sequences). Of the eight most abundant Bacteroides OTUs detected, two were promoted specifically by inulin and six by pectin. Among the Firmicutes, Eubacterium eligens in particular was strongly promoted by pectin, while several species were stimulated by inulin. Responses were influenced by pH, which was stepped up, and down, between 5.5, 6.0, 6.4 and 6.9 in parallel vessels within each experiment. In particular, several experiments involving downshifts to pH 5.5 resulted in Faecalibacterium prausnitzii replacing Bacteroides spp. as the dominant sequences observed. Community diversity was greater in the pectin-fed than in the inulin-fed fermentors, presumably reflecting the differing complexity of the two substrates.
Conclusions: We have shown that particular non-digestible dietary carbohydrates have enormous potential for modifying the gut microbiota, but these modifications occur at the level of individual strains and species and are not easily predicted a priori. Furthermore, the gut environment, especially pH, plays a key role in determining the outcome of interspecies competition. This makes it crucial to put greater effort into identifying the range of bacteria that may be stimulated by a given prebiotic approach. Both for reasons of efficacy and of safety, the development of prebiotics intended to benefit human health has to take account of the highly individual species profiles that may result.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
