

Mass Spectrometry-Based Quantitative Strategies for Assessing the Biological Consequences and Repair of DNA Adducts
- PMID: 26758048
- PMCID: PMC5047065
- DOI: 10.1021/acs.accounts.5b00437
Mass Spectrometry-Based Quantitative Strategies for Assessing the Biological Consequences and Repair of DNA Adducts
Abstract
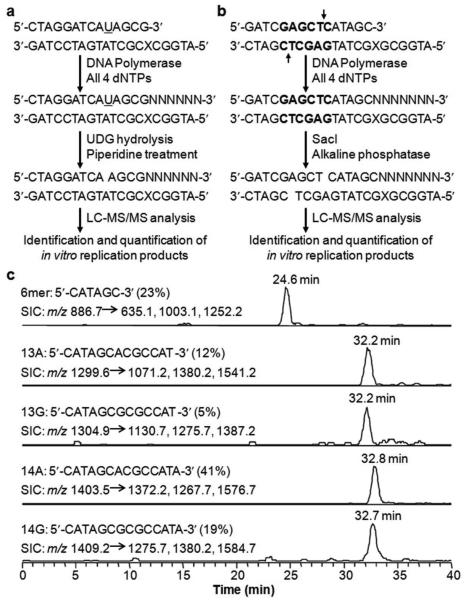
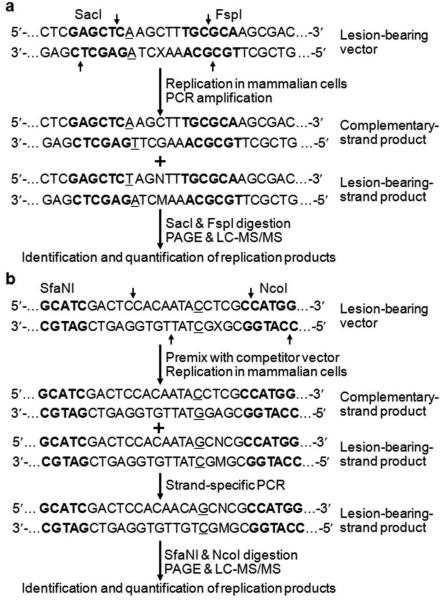
The genetic integrity of living organisms is constantly threatened by environmental and endogenous sources of DNA damaging agents that can induce a plethora of chemically modified DNA lesions. Unrepaired DNA lesions may elicit cytotoxic and mutagenic effects and contribute to the development of human diseases including cancer and neurodegeneration. Understanding the deleterious outcomes of DNA damage necessitates the investigation about the effects of DNA adducts on the efficiency and fidelity of DNA replication and transcription. Conventional methods for measuring lesion-induced replicative or transcriptional alterations often require time-consuming colony screening and DNA sequencing procedures. Recently, a series of mass spectrometry (MS)-based strategies have been developed in our laboratory as an efficient platform for qualitative and quantitative analyses of the changes in genetic information induced by DNA adducts during DNA replication and transcription. During the past few years, we have successfully used these MS-based methods for assessing the replicative or transcriptional blocking and miscoding properties of more than 30 distinct DNA adducts. When combined with genetic manipulation, these methods have also been successfully employed for revealing the roles of various DNA repair proteins or translesion synthesis DNA polymerases (Pols) in modulating the adverse effects of DNA lesions on transcription or replication in mammalian and bacterial cells. For instance, we found that Escherichia coli Pol IV and its mammalian ortholog (i.e., Pol κ) are required for error-free bypass of N(2)-(1-carboxyethyl)-2'-deoxyguanosine (N(2)-CEdG) in cells. We also found that the N(2)-CEdG lesions strongly inhibit DNA transcription and they are repaired by transcription-coupled nucleotide excision repair in mammalian cells. In this Account, we focus on the development of MS-based approaches for determining the effects of DNA adducts on DNA replication and transcription, where liquid chromatography-tandem mass spectrometry is employed for the identification, and sometimes quantification, of the progeny products arising from the replication or transcription of lesion-bearing DNA substrates in vitro and in mammalian cells. We also highlight their applications to lesion bypass, mutagenesis, and repair studies of three representative types of DNA lesions, that is, the methylglyoxal-induced N(2)-CEdG, oxidatively induced 8,5'-cyclopurine-2'-deoxynucleosides, and regioisomeric alkylated thymidine lesions. Specially, we discuss the similar and distinct effects of the minor-groove DNA lesions including N(2)-CEdG and O(2)-alkylated thymidine lesions, as well as the major-groove O(4)-alkylated thymidine lesions on DNA replication and transcription machinery. For example, we found that the addition of an alkyl group to the O(4) position of thymine may facilitate its preferential pairing with guanine and thus induce exclusively the misincorporation of guanine nucleotide opposite the lesion, whereas alkylation of thymine at the O(2) position may render the nucleobase unfavorable in pairing with any of the canonical nucleobases and thus exhibit promiscuous miscoding properties during DNA replication and transcription. The MS-based strategies described herein should be generally applicable for quantitative measurement of the biological consequences and repair of other DNA lesions in vitro and in cells.
Figures








Similar articles
-
Cytotoxic and mutagenic properties of O4-alkylthymidine lesions in Escherichia coli cells.Nucleic Acids Res. 2015 Dec 15;43(22):10795-803. doi: 10.1093/nar/gkv941. Epub 2015 Sep 22. Nucleic Acids Res. 2015. PMID: 26400162 Free PMC article.
-
Repair and translesion synthesis of O6-alkylguanine DNA lesions in human cells.J Biol Chem. 2019 Jul 19;294(29):11144-11153. doi: 10.1074/jbc.RA119.009054. Epub 2019 Jun 5. J Biol Chem. 2019. PMID: 31167778 Free PMC article.
-
Mechanism of DNA alkylation-induced transcriptional stalling, lesion bypass, and mutagenesis.Proc Natl Acad Sci U S A. 2017 Aug 22;114(34):E7082-E7091. doi: 10.1073/pnas.1708748114. Epub 2017 Aug 7. Proc Natl Acad Sci U S A. 2017. PMID: 28784758 Free PMC article.
-
Chemistry and biology of DNA containing 1,N(2)-deoxyguanosine adducts of the alpha,beta-unsaturated aldehydes acrolein, crotonaldehyde, and 4-hydroxynonenal.Chem Res Toxicol. 2009 May;22(5):759-78. doi: 10.1021/tx9000489. Chem Res Toxicol. 2009. PMID: 19397281 Free PMC article. Review.
-
Postreplication repair mechanisms in the presence of DNA adducts in Escherichia coli.Mutat Res. 2011 May-Jun;727(3):104-22. doi: 10.1016/j.mrrev.2011.04.003. Epub 2011 May 5. Mutat Res. 2011. PMID: 21558018 Review.
Cited by
-
The Impact of Minor-Groove N2-Alkyl-2'-deoxyguanosine Lesions on DNA Replication in Human Cells.ACS Chem Biol. 2019 Aug 16;14(8):1708-1716. doi: 10.1021/acschembio.9b00129. Epub 2019 Aug 1. ACS Chem Biol. 2019. PMID: 31347832 Free PMC article.
-
Biological Evaluation of DNA Biomarkers in a Chemically Defined and Site-Specific Manner.Toxics. 2019 Jun 25;7(2):36. doi: 10.3390/toxics7020036. Toxics. 2019. PMID: 31242562 Free PMC article. Review.
-
N 2-Alkyl-dG lesions elicit R-loop accumulation in the genome.Nucleic Acids Res. 2024 Nov 11;52(20):12487-12497. doi: 10.1093/nar/gkae845. Nucleic Acids Res. 2024. PMID: 39351875 Free PMC article.
-
Position-dependent effects of regioisomeric methylated adenine and guanine ribonucleosides on translation.Nucleic Acids Res. 2017 Sep 6;45(15):9059-9067. doi: 10.1093/nar/gkx515. Nucleic Acids Res. 2017. PMID: 28591780 Free PMC article.
-
Roles of Aag, Alkbh2, and Alkbh3 in the Repair of Carboxymethylated and Ethylated Thymidine Lesions.ACS Chem Biol. 2016 May 20;11(5):1332-8. doi: 10.1021/acschembio.6b00085. Epub 2016 Mar 4. ACS Chem Biol. 2016. PMID: 26930515 Free PMC article.
References
-
- Ziv O, Diamant N, Shachar S, Hendel A, Livneh Z. Quantitative measurement of translesion DNA synthesis in mammalian cells. Methods Mol. Biol. 2012;920:529–542. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
