

Molecular and Structural Analysis of the Helicobacter pylori cag Type IV Secretion System Core Complex
- PMID: 26758182
- PMCID: PMC4725015
- DOI: 10.1128/mBio.02001-15
Molecular and Structural Analysis of the Helicobacter pylori cag Type IV Secretion System Core Complex
Abstract
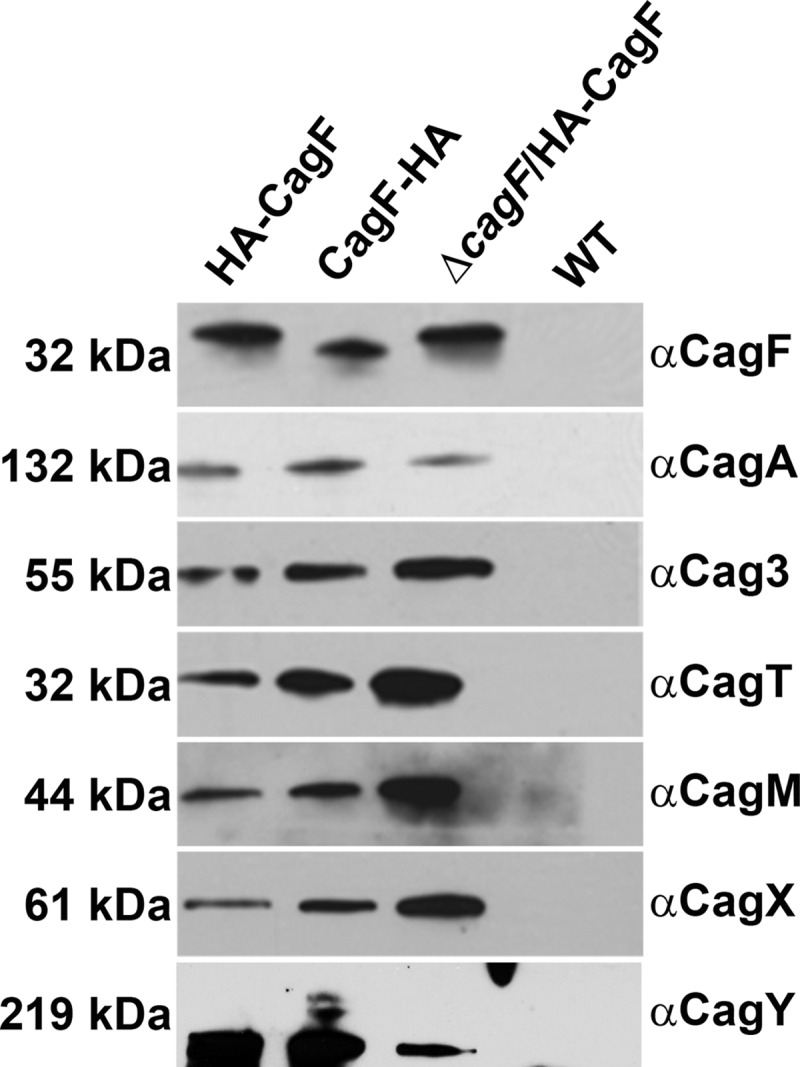
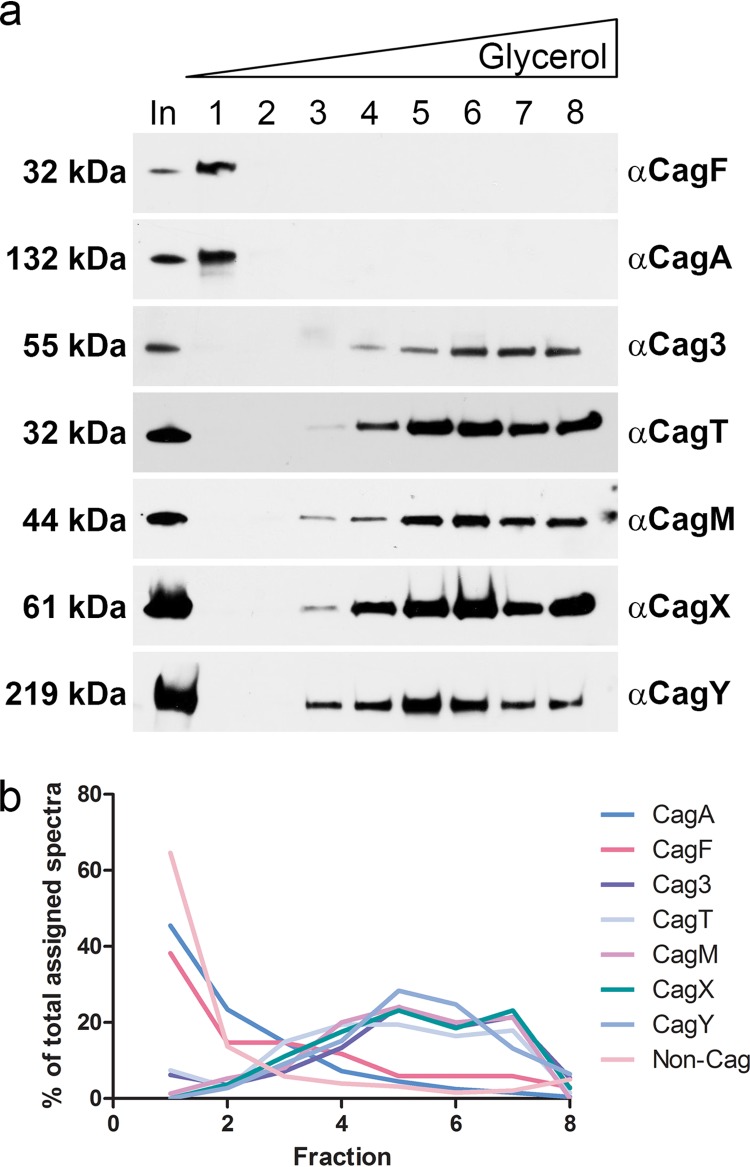
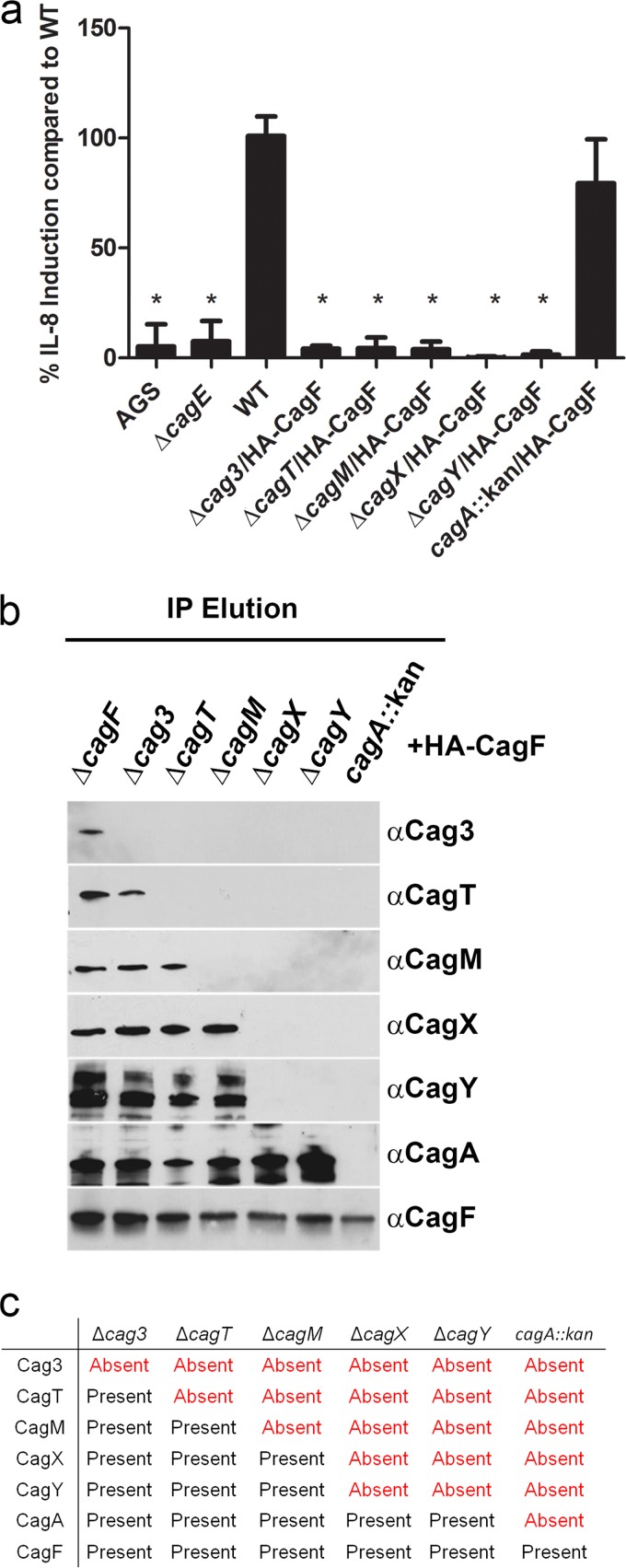
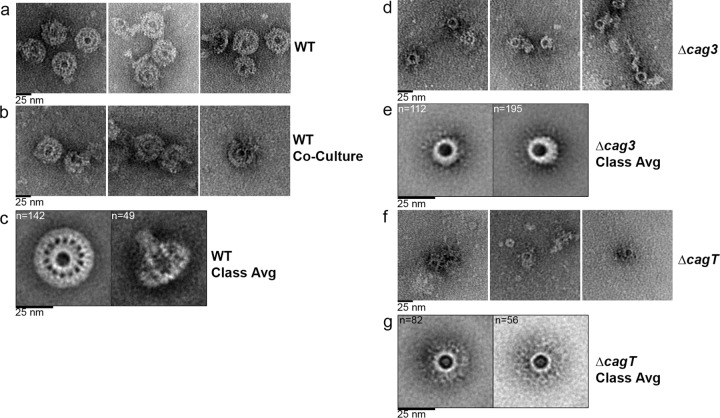
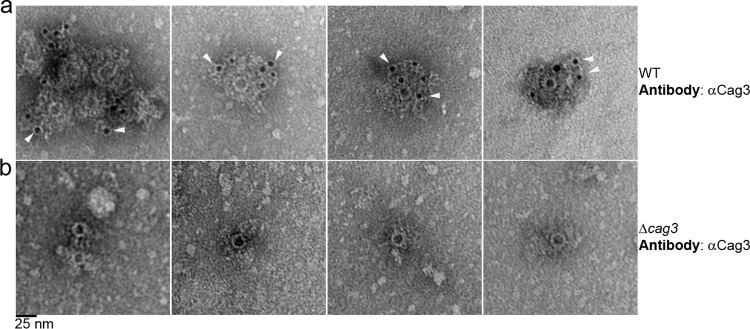
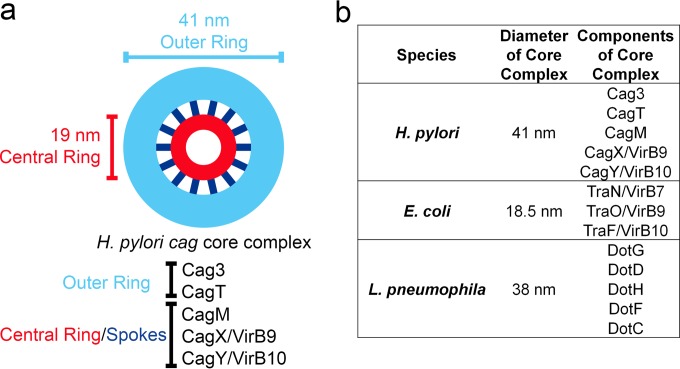
Bacterial type IV secretion systems (T4SSs) can function to export or import DNA, and can deliver effector proteins into a wide range of target cells. Relatively little is known about the structural organization of T4SSs that secrete effector proteins. In this report, we describe the isolation and analysis of a membrane-spanning core complex from the Helicobacter pylori cag T4SS, which has an important role in the pathogenesis of gastric cancer. We show that this complex contains five H. pylori proteins, CagM, CagT, Cag3, CagX, and CagY, each of which is required for cag T4SS activity. CagX and CagY are orthologous to the VirB9 and VirB10 components of T4SSs in other bacterial species, and the other three Cag proteins are unique to H. pylori. Negative stain single-particle electron microscopy revealed complexes 41 nm in diameter, characterized by a 19-nm-diameter central ring linked to an outer ring by spoke-like linkers. Incomplete complexes formed by Δcag3 or ΔcagT mutants retain the 19-nm-diameter ring but lack an organized outer ring. Immunogold labeling studies confirm that Cag3 is a peripheral component of the complex. The cag T4SS core complex has an overall diameter and structural organization that differ considerably from the corresponding features of conjugative T4SSs. These results highlight specialized features of the H. pylori cag T4SS that are optimized for function in the human gastric mucosal environment.
Importance: Type IV secretion systems (T4SSs) are versatile macromolecular machines that are present in many bacterial species. In this study, we investigated a T4SS found in the bacterium Helicobacter pylori. H. pylori is an important cause of stomach cancer, and the H. pylori T4SS contributes to cancer pathogenesis by mediating entry of CagA (an effector protein regarded as a "bacterial oncoprotein") into gastric epithelial cells. We isolated and analyzed the membrane-spanning core complex of the H. pylori T4SS and showed that it contains unique proteins unrelated to components of T4SSs in other bacterial species. These results constitute the first structural analysis of the core complex from this important secretion system.
Copyright © 2016 Frick-Cheng et al.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
