

Arabidopsis Flower and Embryo Developmental Genes are Repressed in Seedlings by Different Combinations of Polycomb Group Proteins in Association with Distinct Sets of Cis-regulatory Elements
- PMID: 26760036
- PMCID: PMC4711971
- DOI: 10.1371/journal.pgen.1005771
Arabidopsis Flower and Embryo Developmental Genes are Repressed in Seedlings by Different Combinations of Polycomb Group Proteins in Association with Distinct Sets of Cis-regulatory Elements
Erratum in
-
Correction: Arabidopsis Flower and Embryo Developmental Genes are Repressed in Seedlings by Different Combinations of Polycomb Group Proteins in Association with Distinct Sets of Cis-regulatory Elements.PLoS Genet. 2017 Jan 25;13(1):e1006574. doi: 10.1371/journal.pgen.1006574. eCollection 2017 Jan. PLoS Genet. 2017. PMID: 28122004 Free PMC article.
Abstract
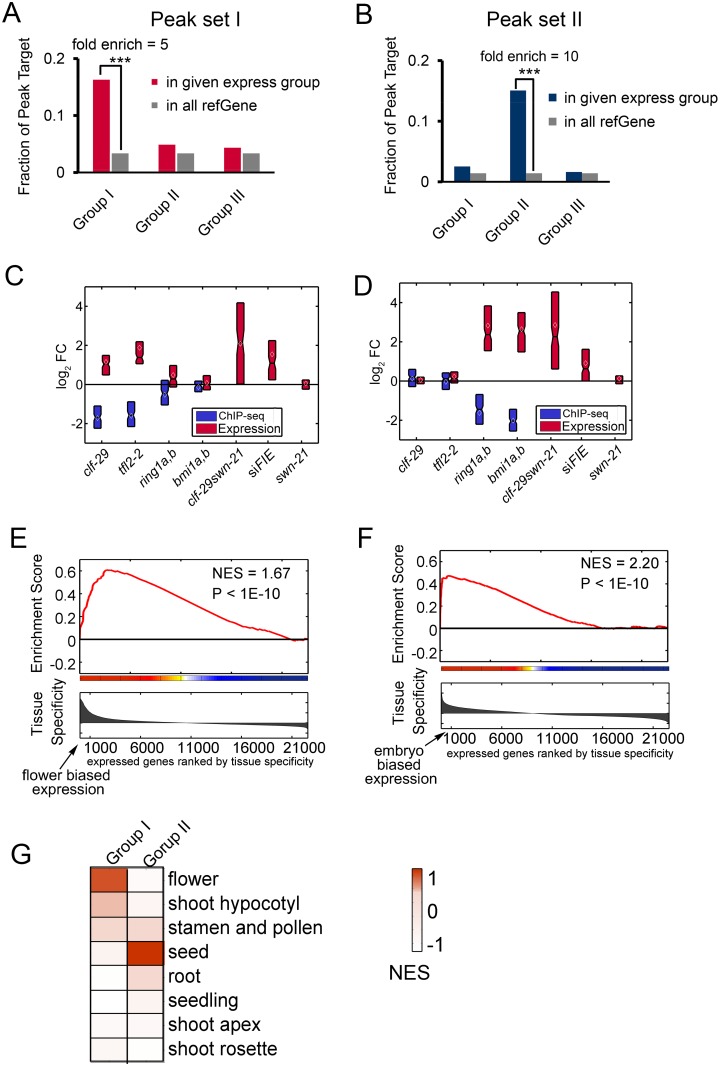
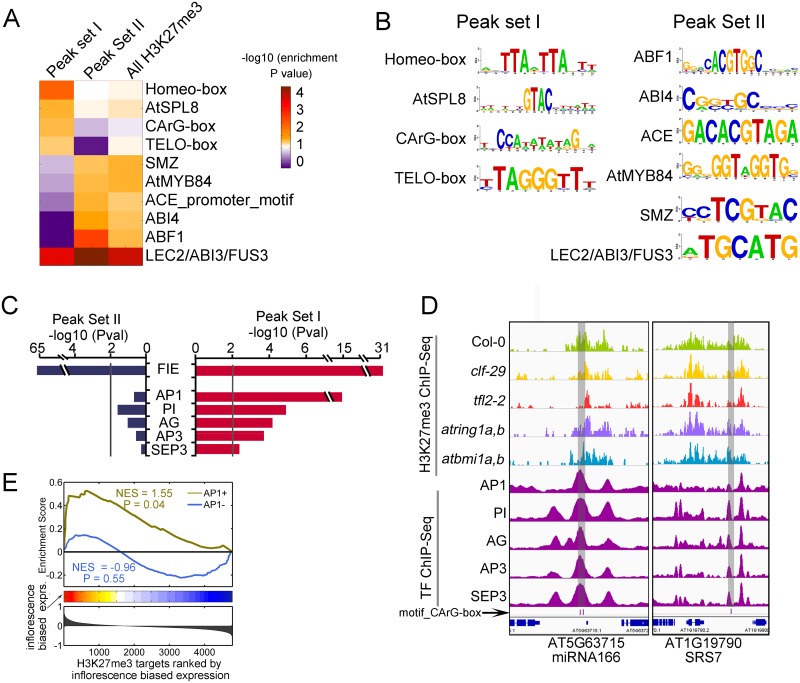
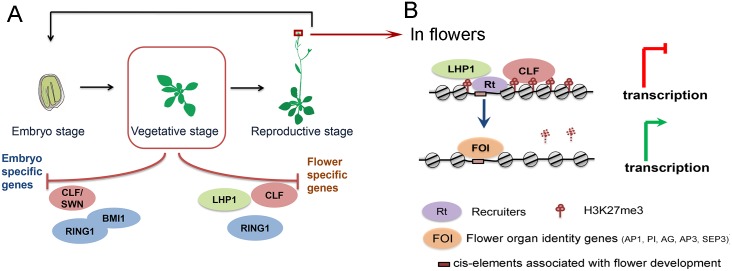
Polycomb repressive complexes (PRCs) play crucial roles in transcriptional repression and developmental regulation in both plants and animals. In plants, depletion of different members of PRCs causes both overlapping and unique phenotypic defects. However, the underlying molecular mechanism determining the target specificity and functional diversity is not sufficiently characterized. Here, we quantitatively compared changes of tri-methylation at H3K27 in Arabidopsis mutants deprived of various key PRC components. We show that CURLY LEAF (CLF), a major catalytic subunit of PRC2, coordinates with different members of PRC1 in suppression of distinct plant developmental programs. We found that expression of flower development genes is repressed in seedlings preferentially via non-redundant role of CLF, which specifically associated with LIKE HETEROCHROMATIN PROTEIN1 (LHP1). In contrast, expression of embryo development genes is repressed by PRC1-catalytic core subunits AtBMI1 and AtRING1 in common with PRC2-catalytic enzymes CLF or SWINGER (SWN). This context-dependent role of CLF corresponds well with the change in H3K27me3 profiles, and is remarkably associated with differential co-occupancy of binding motifs of transcription factors (TFs), including MADS box and ABA-related factors. We propose that different combinations of PRC members distinctively regulate different developmental programs, and their target specificity is modulated by specific TFs.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Mozgova I, Hennig L (2015) The Polycomb Group Protein Regulatory Network. Annu Rev Plant Biol. - PubMed
-
- Xiao J, Wagner D (2015) Polycomb repression in the regulation of growth and development in Arabidopsis. Curr Opin Plant Biol 23C: 15–24. - PubMed
-
- Goodrich J, Puangsomlee P, Martin M, Long D, Meyerowitz EM, et al. (1997) A Polycomb-group gene regulates homeotic gene expression in Arabidopsis. Nature 386: 44–51. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
