

The Roles of RNase-L in Antimicrobial Immunity and the Cytoskeleton-Associated Innate Response
- PMID: 26760998
- PMCID: PMC4730318
- DOI: 10.3390/ijms17010074
The Roles of RNase-L in Antimicrobial Immunity and the Cytoskeleton-Associated Innate Response
Abstract
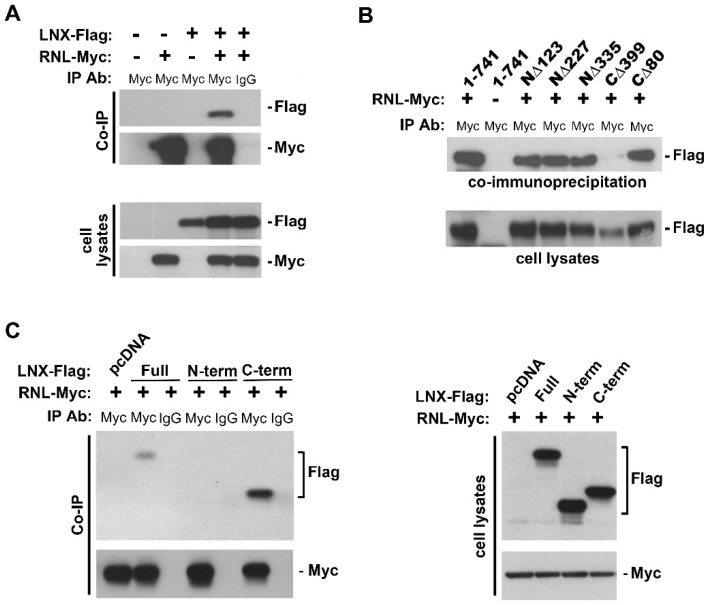
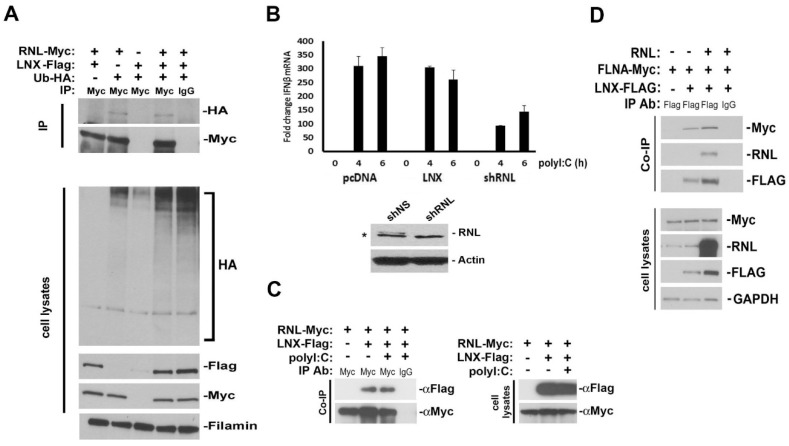
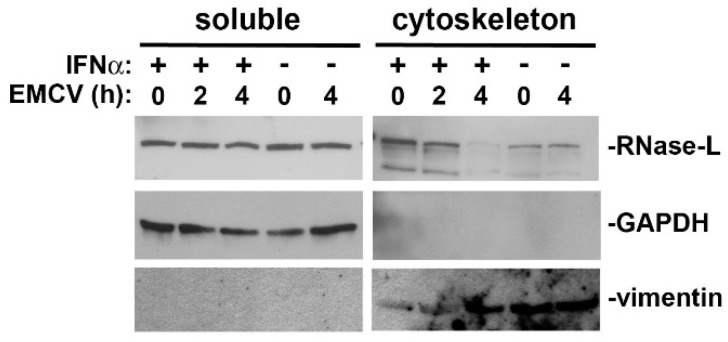
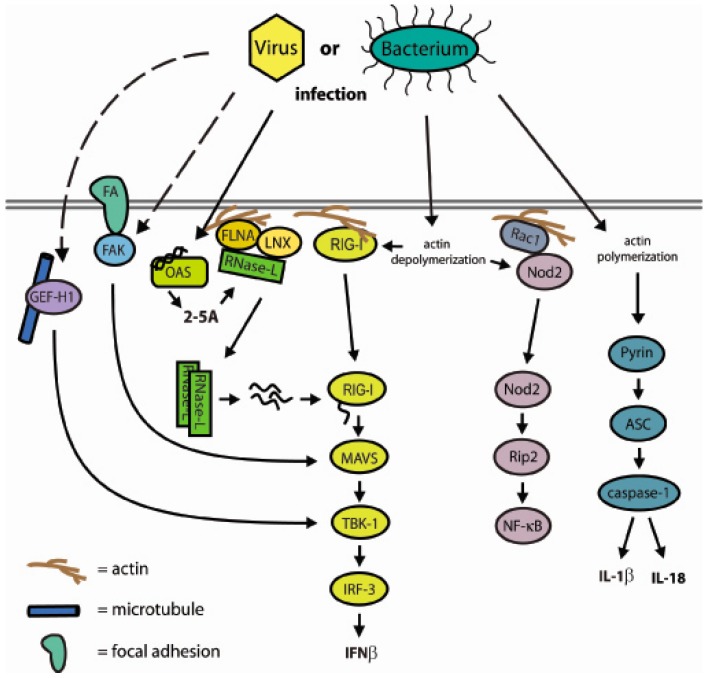
The interferon (IFN)-regulated endoribonuclease RNase-L is involved in multiple aspects of the antimicrobial innate immune response. It is the terminal component of an RNA cleavage pathway in which dsRNA induces the production of RNase-L-activating 2-5A by the 2'-5'-oligoadenylate synthetase. The active nuclease then cleaves ssRNAs, both cellular and viral, leading to downregulation of their expression and the generation of small RNAs capable of activating retinoic acid-inducible gene-I (RIG-I)-like receptors or the nucleotide-binding oligomerization domain-like receptor 3 (NLRP3) inflammasome. This leads to IFNβ expression and IL-1β activation respectively, in addition to broader effects on immune cell function. RNase-L is also one of a growing number of innate immune components that interact with the cell cytoskeleton. It can bind to several cytoskeletal proteins, including filamin A, an actin-binding protein that collaborates with RNase-L to maintain the cellular barrier to viral entry. This antiviral activity is independent of catalytic function, a unique mechanism for RNase-L. We also describe here the interaction of RNase-L with the E3 ubiquitin ligase and scaffolding protein, ligand of nump protein X (LNX), a regulator of tight junction proteins. In order to better understand the significance and context of these novel binding partners in the antimicrobial response, other innate immune protein interactions with the cytoskeleton are also discussed.
Keywords: LNX; RNase-L; actin; cytoskeleton; filamin A; inflammasome; innate immunity; interferon.
Figures

References
-
- Brand S., Zitzmann K., Dambacher J., Beigel F., Olszak T., Vlotides G., Eichhorst S.T., Goke B., Diepolder H., Auernhammer C.J. SOCS-1 inhibits expression of the antiviral proteins 2′,5′-OAS and MxA induced by the novel interferon-λs IL-28a and IL-29. Biochem. Biophys. Res. Commun. 2005;331:543–548. doi: 10.1016/j.bbrc.2005.04.004. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
