

Asymmetrical Damage Partitioning in Bacteria: A Model for the Evolution of Stochasticity, Determinism, and Genetic Assimilation
- PMID: 26761487
- PMCID: PMC4711911
- DOI: 10.1371/journal.pcbi.1004700
Asymmetrical Damage Partitioning in Bacteria: A Model for the Evolution of Stochasticity, Determinism, and Genetic Assimilation
Abstract
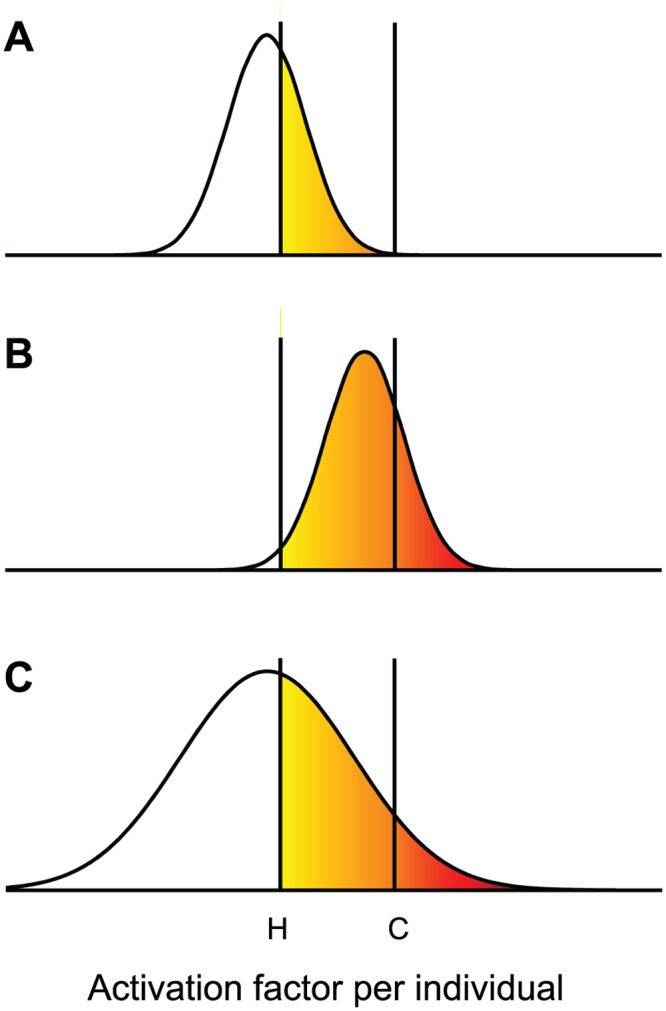
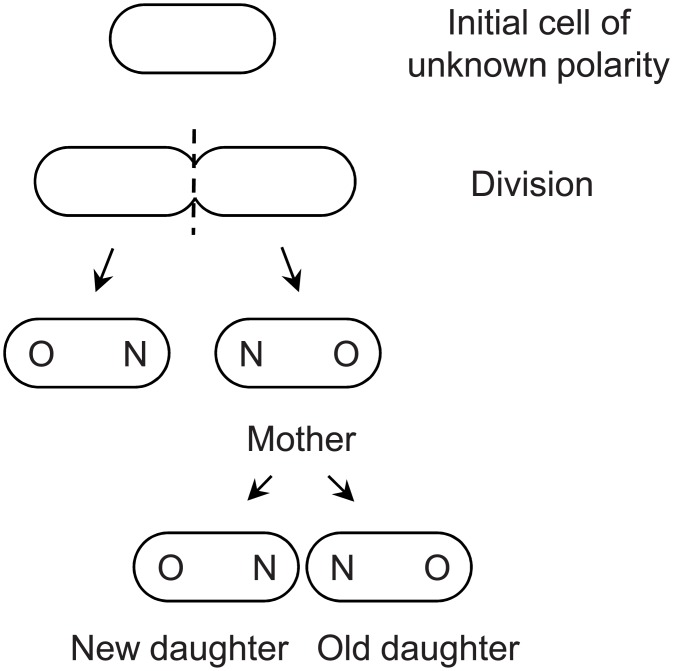
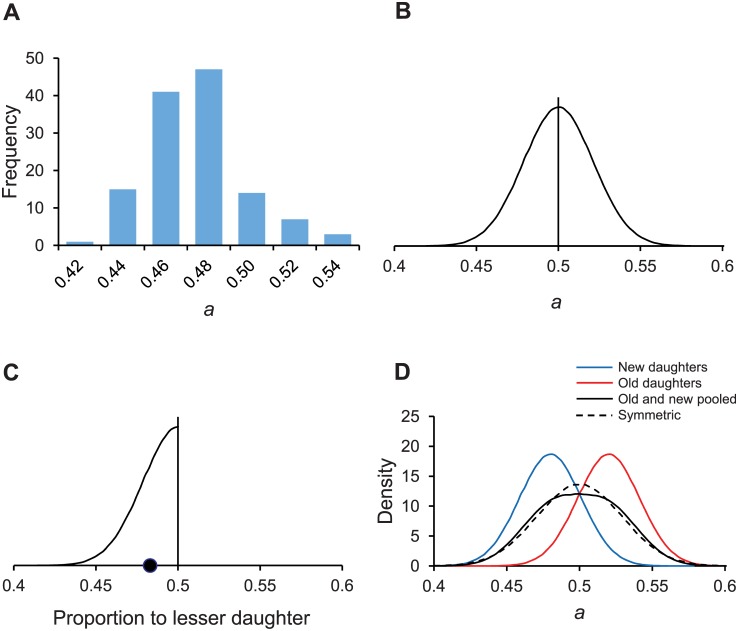
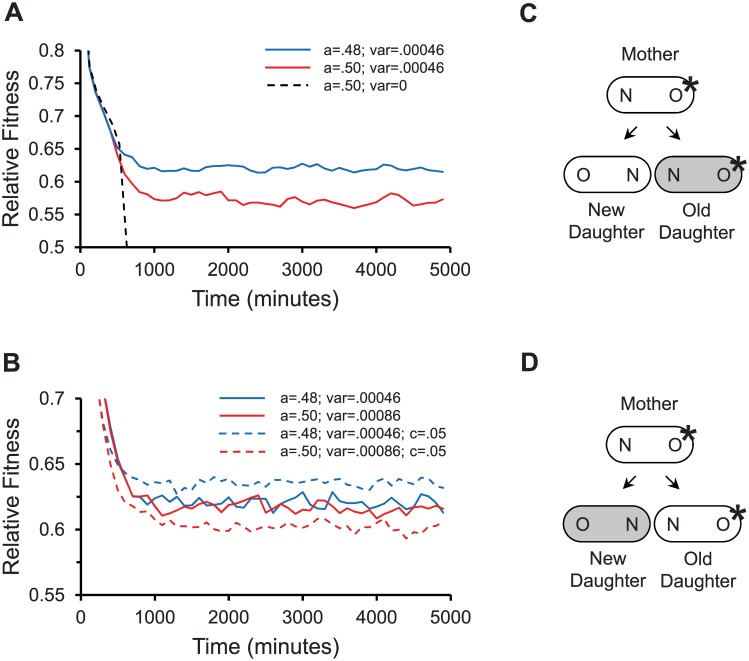
Non-genetic phenotypic variation is common in biological organisms. The variation is potentially beneficial if the environment is changing. If the benefit is large, selection can favor the evolution of genetic assimilation, the process by which the expression of a trait is transferred from environmental to genetic control. Genetic assimilation is an important evolutionary transition, but it is poorly understood because the fitness costs and benefits of variation are often unknown. Here we show that the partitioning of damage by a mother bacterium to its two daughters can evolve through genetic assimilation. Bacterial phenotypes are also highly variable. Because gene-regulating elements can have low copy numbers, the variation is attributed to stochastic sampling. Extant Escherichia coli partition asymmetrically and deterministically more damage to the old daughter, the one receiving the mother's old pole. By modeling in silico damage partitioning in a population, we show that deterministic asymmetry is advantageous because it increases fitness variance and hence the efficiency of natural selection. However, we find that symmetrical but stochastic partitioning can be similarly beneficial. To examine why bacteria evolved deterministic asymmetry, we modeled the effect of damage anchored to the mother's old pole. While anchored damage strengthens selection for asymmetry by creating additional fitness variance, it has the opposite effect on symmetry. The difference results because anchored damage reinforces the polarization of partitioning in asymmetric bacteria. In symmetric bacteria, it dilutes the polarization. Thus, stochasticity alone may have protected early bacteria from damage, but deterministic asymmetry has evolved to be equally important in extant bacteria. We estimate that 47% of damage partitioning is deterministic in E. coli. We suggest that the evolution of deterministic asymmetry from stochasticity offers an example of Waddington's genetic assimilation. Our model is able to quantify the evolution of the assimilation because it characterizes the fitness consequences of variation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Allocation of gene products to daughter cells is determined by the age of the mother in single Escherichia coli cells.Proc Biol Sci. 2020 May 13;287(1926):20200569. doi: 10.1098/rspb.2020.0569. Epub 2020 May 6. Proc Biol Sci. 2020. PMID: 32370668 Free PMC article.
-
A model for damage load and its implications for the evolution of bacterial aging.PLoS Genet. 2010 Aug 26;6(8):e1001076. doi: 10.1371/journal.pgen.1001076. PLoS Genet. 2010. PMID: 20865171 Free PMC article.
-
Beyond the (Geometric) Mean: Stochastic Models Undermine Deterministic Predictions of Bet Hedger Evolution.Am Nat. 2025 Jun;205(6):572-589. doi: 10.1086/735690. Epub 2025 May 1. Am Nat. 2025. PMID: 40446303
-
The concept of fitness in fluctuating environments.Trends Ecol Evol. 2015 May;30(5):273-81. doi: 10.1016/j.tree.2015.03.007. Epub 2015 Apr 3. Trends Ecol Evol. 2015. PMID: 25843273 Review.
-
Stochasticity in evolution.Trends Ecol Evol. 2009 Mar;24(3):157-65. doi: 10.1016/j.tree.2008.09.014. Epub 2009 Jan 27. Trends Ecol Evol. 2009. PMID: 19178980 Review.
Cited by
-
Age structure landscapes emerge from the equilibrium between aging and rejuvenation in bacterial populations.Nat Commun. 2018 Sep 13;9(1):3722. doi: 10.1038/s41467-018-06154-9. Nat Commun. 2018. PMID: 30213942 Free PMC article.
-
Bacterial ageing in the absence of external stressors.Philos Trans R Soc Lond B Biol Sci. 2019 Nov 25;374(1786):20180442. doi: 10.1098/rstb.2018.0442. Epub 2019 Oct 7. Philos Trans R Soc Lond B Biol Sci. 2019. PMID: 31587633 Free PMC article.
-
The interplay of phenotypic variability and fitness in finite microbial populations.J R Soc Interface. 2020 May;17(166):20190827. doi: 10.1098/rsif.2019.0827. Epub 2020 May 13. J R Soc Interface. 2020. PMID: 32396808 Free PMC article.
-
Protein aggregation in bacteria.FEMS Microbiol Rev. 2020 Jan 1;44(1):54-72. doi: 10.1093/femsre/fuz026. FEMS Microbiol Rev. 2020. PMID: 31633151 Free PMC article. Review.
-
A Tale of Two Concepts: Harmonizing the Free Radical and Antagonistic Pleiotropy Theories of Aging.Antioxid Redox Signal. 2018 Oct 1;29(10):1003-1017. doi: 10.1089/ars.2017.7105. Epub 2017 Oct 17. Antioxid Redox Signal. 2018. PMID: 28874059 Free PMC article. Review.
References
-
- Falconer DS. Introduction to Quantitative Genetics. 2nd ed New York, N.Y.: Longman Inc.; 1981.
-
- Pigliucci M, Murren CJ, Schlichting CD. Phenotypic plasticity and evolution by genetic assimilation. Journal of Experimental Biology. 2006. June 15; 209(12):2362–7. - PubMed
-
- Schmalhausen II. Factors of evolution: The Theory of Stabilizing Selection Originally Published: Blakiston Co., Philadelphia; 1949. Reprint edition: University of Chicago Press, Chicago; 1986.
-
- Waddington CH. Canalization of development and the inheritance of acquired characters. Nature. 1942. Jul-Dec; 150:563–5. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
