

Recruitment of the 4EHP-GYF2 cap-binding complex to tetraproline motifs of tristetraprolin promotes repression and degradation of mRNAs with AU-rich elements
- PMID: 26763119
- PMCID: PMC4748815
- DOI: 10.1261/rna.054833.115
Recruitment of the 4EHP-GYF2 cap-binding complex to tetraproline motifs of tristetraprolin promotes repression and degradation of mRNAs with AU-rich elements
Abstract
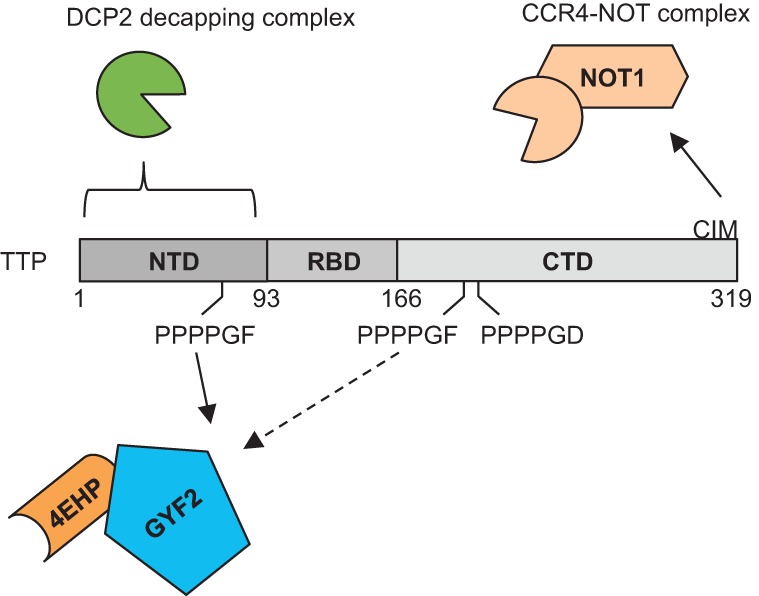
The zinc finger protein tristetraprolin (TTP) promotes translation repression and degradation of mRNAs containing AU-rich elements (AREs). Although much attention has been directed toward understanding the decay process and machinery involved, the translation repression role of TTP has remained poorly understood. Here we identify the cap-binding translation repression 4EHP-GYF2 complex as a cofactor of TTP. Immunoprecipitation and in vitro pull-down assays demonstrate that TTP associates with the 4EHP-GYF2 complex via direct interaction with GYF2, and mutational analyses show that this interaction occurs via conserved tetraproline motifs of TTP. Mutant TTP with diminished 4EHP-GYF2 binding is impaired in its ability to repress a luciferase reporter ARE-mRNA. 4EHP knockout mouse embryonic fibroblasts (MEFs) display increased induction and slower turnover of TTP-target mRNAs as compared to wild-type MEFs. Our work highlights the function of the conserved tetraproline motifs of TTP and identifies 4EHP-GYF2 as a cofactor in translational repression and mRNA decay by TTP.
Keywords: ARE-mediated decay; AU-rich elements; mRNA turnover; translation repression; tristetraprolin.
© 2016 Fu et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures

Similar articles
-
Tristetraprolin Recruits Eukaryotic Initiation Factor 4E2 To Repress Translation of AU-Rich Element-Containing mRNAs.Mol Cell Biol. 2015 Nov;35(22):3921-32. doi: 10.1128/MCB.00845-15. Epub 2015 Sep 14. Mol Cell Biol. 2015. PMID: 26370510 Free PMC article.
-
Direct role for the Drosophila GIGYF protein in 4EHP-mediated mRNA repression.Nucleic Acids Res. 2019 Jul 26;47(13):7035-7048. doi: 10.1093/nar/gkz429. Nucleic Acids Res. 2019. PMID: 31114929 Free PMC article.
-
Tristetraprolin Recruits the Herpes Simplex Virion Host Shutoff RNase to AU-Rich Elements in Stress Response mRNAs To Enable Their Cleavage.J Virol. 2015 May;89(10):5643-50. doi: 10.1128/JVI.00091-15. Epub 2015 Mar 11. J Virol. 2015. PMID: 25762736 Free PMC article.
-
The Cross-talk between Tristetraprolin and Cytokines in Cancer.Anticancer Agents Med Chem. 2017 Nov 24;17(11):1477-1486. doi: 10.2174/1871520617666170327155124. Anticancer Agents Med Chem. 2017. PMID: 28356023 Review.
-
The role of tristetraprolin in cancer and inflammation.Front Biosci (Landmark Ed). 2012 Jan 1;17(1):174-88. doi: 10.2741/3920. Front Biosci (Landmark Ed). 2012. PMID: 22201737 Free PMC article. Review.
Cited by
-
FMRP-dependent production of large dosage-sensitive proteins is highly conserved.Genetics. 2022 Jul 30;221(4):iyac094. doi: 10.1093/genetics/iyac094. Genetics. 2022. PMID: 35731217 Free PMC article.
-
GIGYF1/2 proteins use auxiliary sequences to selectively bind to 4EHP and repress target mRNA expression.Genes Dev. 2017 Jun 1;31(11):1147-1161. doi: 10.1101/gad.299420.117. Epub 2017 Jul 11. Genes Dev. 2017. PMID: 28698298 Free PMC article.
-
4EHP and GIGYF1/2 Mediate Translation-Coupled Messenger RNA Decay.Cell Rep. 2020 Oct 13;33(2):108262. doi: 10.1016/j.celrep.2020.108262. Cell Rep. 2020. PMID: 33053355 Free PMC article.
-
GIGYF2 and 4EHP Inhibit Translation Initiation of Defective Messenger RNAs to Assist Ribosome-Associated Quality Control.Mol Cell. 2020 Sep 17;79(6):950-962.e6. doi: 10.1016/j.molcel.2020.07.007. Epub 2020 Jul 28. Mol Cell. 2020. PMID: 32726578 Free PMC article.
-
Major splice variants and multiple polyadenylation site utilization in mRNAs encoding human translation initiation factors eIF4E1 and eIF4E3 regulate the translational regulators?Mol Genet Genomics. 2018 Feb;293(1):167-186. doi: 10.1007/s00438-017-1375-4. Epub 2017 Sep 23. Mol Genet Genomics. 2018. PMID: 28942592
References
-
- Carballo E, Lai WS, Blackshear PJ. 1998. Feedback inhibition of macrophage tumor necrosis factor-α production by tristetraprolin. Science 281: 1001–1005. - PubMed
-
- Carballo E, Lai WS, Blackshear PJ. 2000. Evidence that tristetraprolin is a physiological regulator of granulocyte-macrophage colony-stimulating factor messenger RNA deadenylation and stability. Blood 95: 1891–1899. - PubMed
-
- Chen CY, Gherzi R, Ong SE, Chan EL, Raijmakers R, Pruijn GJ, Stoecklin G, Moroni C, Mann M, Karin M. 2001. AU binding proteins recruit the exosome to degrade ARE-containing mRNAs. Cell 107: 451–464. - PubMed
-
- Cho PF, Poulin F, Cho-Park YA, Cho-Park IB, Chicoine JD, Lasko P, Sonenberg N. 2005. A new paradigm for translational control: inhibition via 5′-3′ mRNA tethering by Bicoid and the eIF4E cognate 4EHP. Cell 121: 411–423. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous