

Comment
doi: 10.1016/j.cub.2015.11.011.
Neuroscience: A Distributed Neural Network Controls REM Sleep
Affiliations
- PMID: 26766231
- PMCID: PMC5846126
- DOI: 10.1016/j.cub.2015.11.011
Item in Clipboard
Comment
Neuroscience: A Distributed Neural Network Controls REM Sleep
Curr Biol.
.
Abstract
How does the brain control dreams? New science shows that a small node of cells in the medulla - the most primitive part of the brain - may function to control REM sleep, the brain state that underlies dreaming.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
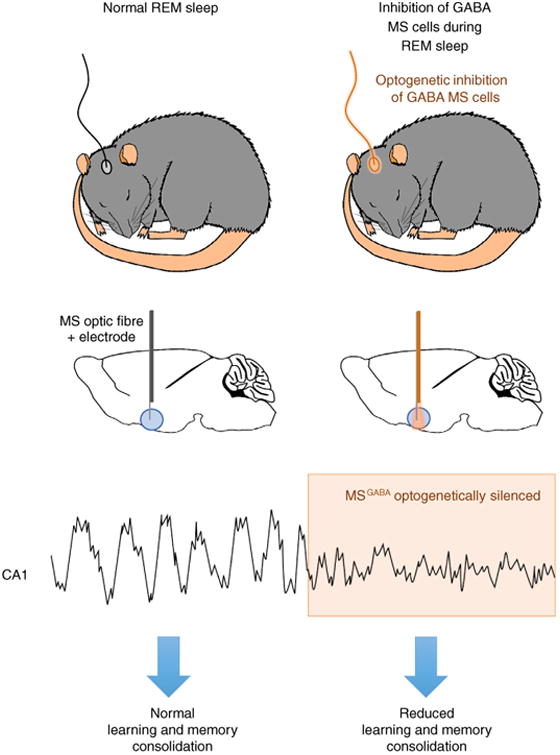
A schematic diagram demonstrating how optogenetic inhibition of GABA cells in the medial septum (MS) of mice affects acquisition of new memory associations. Top two figures show that naturally sleeping mice were fitted with optic fibers targeted above the MS. Delivery of orange laser light to the MS during REM sleep periods allowed for high precision optical inhibition of GABA cells in the MS since these cells expressed the inhibitory opsin archaerhodopsin (ArchT). The panel below the top two shows a sample recording of how optical inhibition of GABA inhibition during REM sleep impacts theta activity. Upon light-induced optogenetic inhibition (orange shading), a rapid reduction in amplitude of hippocampal theta rhythm is observed. REM sleep-specific inhibition of theta activity results in a reduction in learning and memory consolidation.
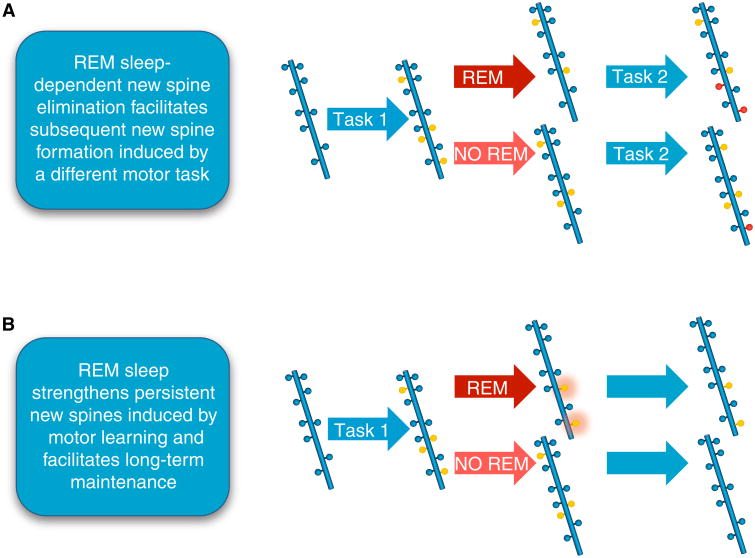
A schematic diagram demonstrating that REM sleep strengthens persistent new spines induced by motor learning and facilitates long-term maintenance of synapses. (A) REM sleep prunes newly formed postsynaptic dendritic spines (yellow circles) of pyramidal neurons in the mouse motor cortex after motor learning (forward walking: Task 1). This REM sleep-dependent elimination of new spines facilitates subsequent spine formation (red circles) when a new motor task is learned (backward walking: Task 2), indicating a role for REM sleep in pruning to balance the number of new spines formed. (B) REM sleep also strengthens persistent new spines (orange halo around yellow circles) induced by motor learning and facilitates long-term maintenance, which are important for behavioral improvement after learning.
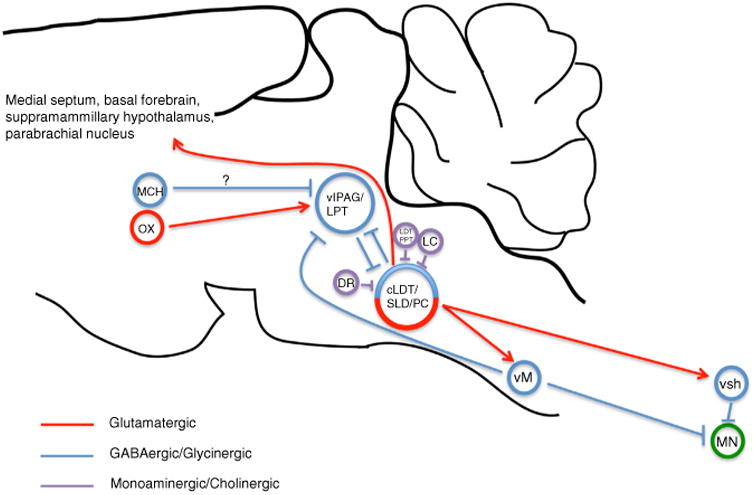
Together the caudal laterodorsal tegmental nucleus (cLDT), sublaterodorsal nucleus (SLD) and precoeruleus region (PC) comprise an executive pontine circuit element for REM sleep. REM-on glutamatergic neurons of the ventral SLD mediate REM motor atonia through direct synaptic activation of glycinergic interneurons of the spinal ventral horn (svh) as well as via GABAergic/glycinergic neurons of the ventral medial medulla (vM). vM neurons in turn project and inhibit motor neurons (MN) of the spinal ventral horn. REM-on GABAergic neurons of the SLD are reciprocally connected with REM-off GABAergic neurons of the ventrolateral periaqueductal gray and lateral pontine tegmentum (vlPAG/LPT), forming the basis of a pontine REM ‘flip-flop’ switch. GABAergic neurons of the vM can also drive REM sleep and do so by inhibiting REM-off vlPAG/LPT neurons. Lateral hypothalamic neurons containing orexin (OX) provide excitatory and stabilizing synaptic control over REM-off vlPAG/LPT neurons. REM-off noradrenergic locus coeruleus (LC) and serotoninergic dorsal raphe (DR) neurons provide additional synaptic control over SLD activity. Cholinergic laterodorsal tegmental and pedunculopontine tegmental (LDT/PPT) neurons may produce REM sleep through activation of REM-on SLD neurons. Lateral hypothalamic neurons containing melanin-concentrating hormone (MCH) also regulate REM sleep, possibly through direct inhibition of REM-off vlPAG/LPT neurons. Glutamatergic neurons of the SLD/PC regulate the corticohippocampal features of REM sleep, likely via projections to the medial septum, parabrachial nucleus, basal forebrain and supramamillary hypothalamus.
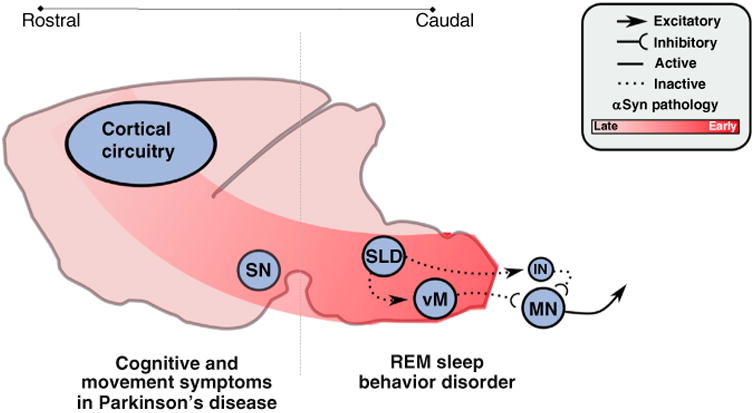
The hypothesized mechanisms and pathways by which synucleinopathic degeneration causes RBD. Synucleinopathic-mediated degeneration begins in the caudal brainstem and spreads inexorably rostrally, in a cell-to-cell fashion. Degeneration initially targets the sublaterodorsal nucleus (SLD) and ventral medulla (vM), impairing their functions and thereby causing loss of REM sleep atonia and the motor behaviors associated with RBD. But, as degenerative processes advance, they spread rostrally into the forebrain circuits that control normal cognition and waking motor behaviors (i.e., the substantia nigra, SN), thus leading to the classic cognitive and motor symptoms associated with synucleinopathies (e.g., Parkinson's disease) that follow initial RBD symptoms.
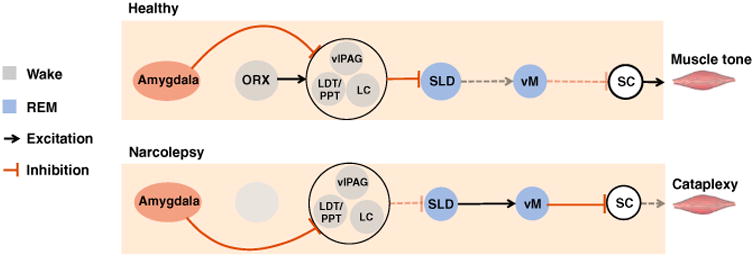
The sublaterodorsal nucleus (SLD) and ventral medulla (vM) represent the core circuits that produce REM sleep atonia. Glutamate cells in the SLD activate GABA and glycine neurons in the vM to trigger REM sleep atonia by inhibiting motoneurons in the spinal cord (SC). In narcolepsy, positive emotions activate GABA neurons in the central nucleus (CeA) that inhibit cells in the locus coeruleus (LC), laterodorsal tegmentum (LDT), and ventrolateral periaqueductal grey (vlPAG), which in turn disinhibit the SLD, which in turn activates the vM to produce motoneuron inhibition and muscle atonia during cataplexy. In non-narcoleptics, positive emotions are unable to trigger muscle atonia because CeA-mediated inhibition of the LC/LDT/vlPAG is offset by excitatory orexin inputs, which prevent positive emotions from manipulating the SLD.
Comment on
-
Control of REM sleep by ventral medulla GABAergic neurons.Nature. 2015 Oct 15;526(7573):435-8. doi: 10.1038/nature14979. Epub 2015 Oct 7. Nature. 2015. PMID: 26444238 Free PMC article.
References
-
- Aserinsky E, Kleitman N. Regularly occurring periods of eye motility, and concomitant phenomena, during sleep. Science. 1953;118:273–274. - PubMed
-
- Dement W, Kleitman N. Cyclic variations in EEG during sleep and their relation to eye movements, body motility, and dreaming. Electroencephalogr Clin Neurophysiol. 1957;9:673–690. - PubMed
-
- Dement W, Kleitman N. The relation of eye movements during sleep to dream activity: an objective method for the study of dreaming. J Exp Psychol. 1957;53:339–346. - PubMed
-
- Jouvet M, Michel F. Electromyographic correlations of sleep in the chronic decorticate & mesencephalic cat. Comptes rendus des seances de la Societe de biologie et de ses filiales. 1959;153:422–425. - PubMed
-
- Jouvet M. The states of sleep. Sci Am. 1967;216:62–68. passim. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
