

Force Feedback Controls Motor Activity and Mechanical Properties of Self-Assembling Branched Actin Networks
- PMID: 26771487
- PMCID: PMC5033619
- DOI: 10.1016/j.cell.2015.11.057
Force Feedback Controls Motor Activity and Mechanical Properties of Self-Assembling Branched Actin Networks
Abstract
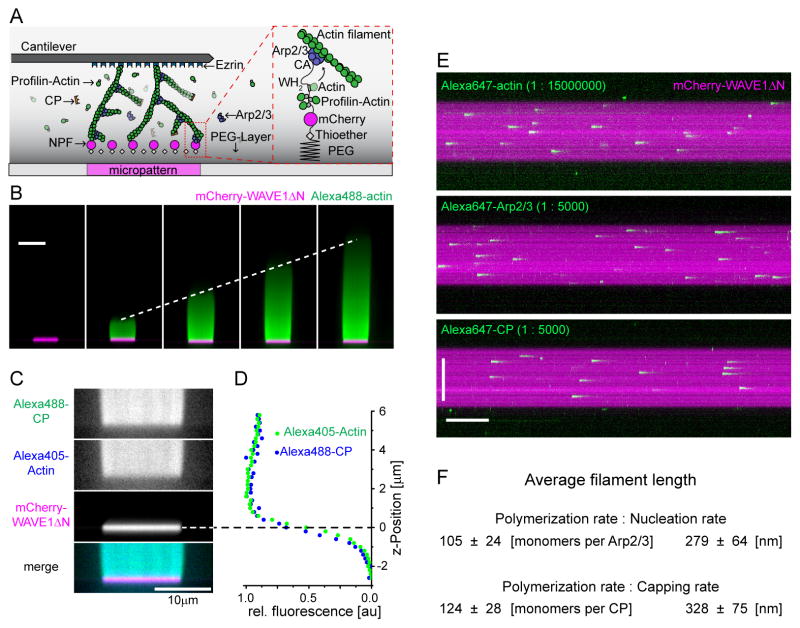
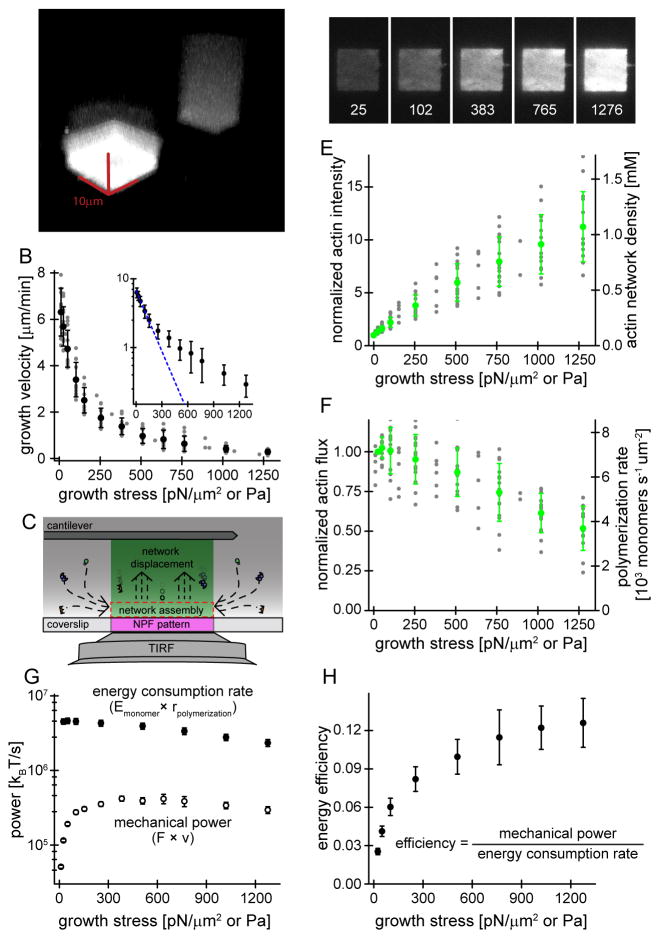
Branched actin networks--created by the Arp2/3 complex, capping protein, and a nucleation promoting factor--generate and transmit forces required for many cellular processes, but their response to force is poorly understood. To address this, we assembled branched actin networks in vitro from purified components and used simultaneous fluorescence and atomic force microscopy to quantify their molecular composition and material properties under various forces. Remarkably, mechanical loading of these self-assembling materials increases their density, power, and efficiency. Microscopically, increased density reflects increased filament number and altered geometry but no change in average length. Macroscopically, increased density enhances network stiffness and resistance to mechanical failure beyond those of isotropic actin networks. These effects endow branched actin networks with memory of their mechanical history that shapes their material properties and motor activity. This work reveals intrinsic force feedback mechanisms by which mechanical resistance makes self-assembling actin networks stiffer, stronger, and more powerful.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures





References
-
- Beltzner CC, Pollard TD. Pathway of actin filament branch formation by Arp2/3 complex. J Biol Chem. 2008;283:7135–7144. - PubMed
-
- Bisi S, Disanza A, Malinverno C, Frittoli E, Palamidessi A, Scita G. Membrane and actin dynamics interplay at lamellipodia leading edge. Curr Opin Cell Biol. 2013;25:565–573. - PubMed
-
- Cameron LA, Svitkina TM, Vignjevic D, Theriot JA, Borisy GG. Dendritic organization of actin comet tails. Curr Biol. 2001;11:130–135. - PubMed
-
- Carter NJ, Cross RA. Mechanics of the kinesin step. Nature. 2005;435:308–312. - PubMed
