

Restoration of Visual Function by Enhancing Conduction in Regenerated Axons
- PMID: 26771493
- PMCID: PMC4863988
- DOI: 10.1016/j.cell.2015.11.036
Restoration of Visual Function by Enhancing Conduction in Regenerated Axons
Abstract
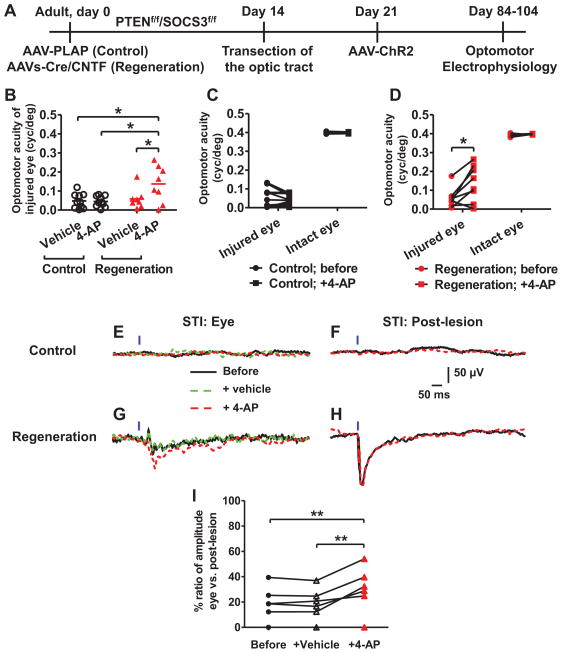
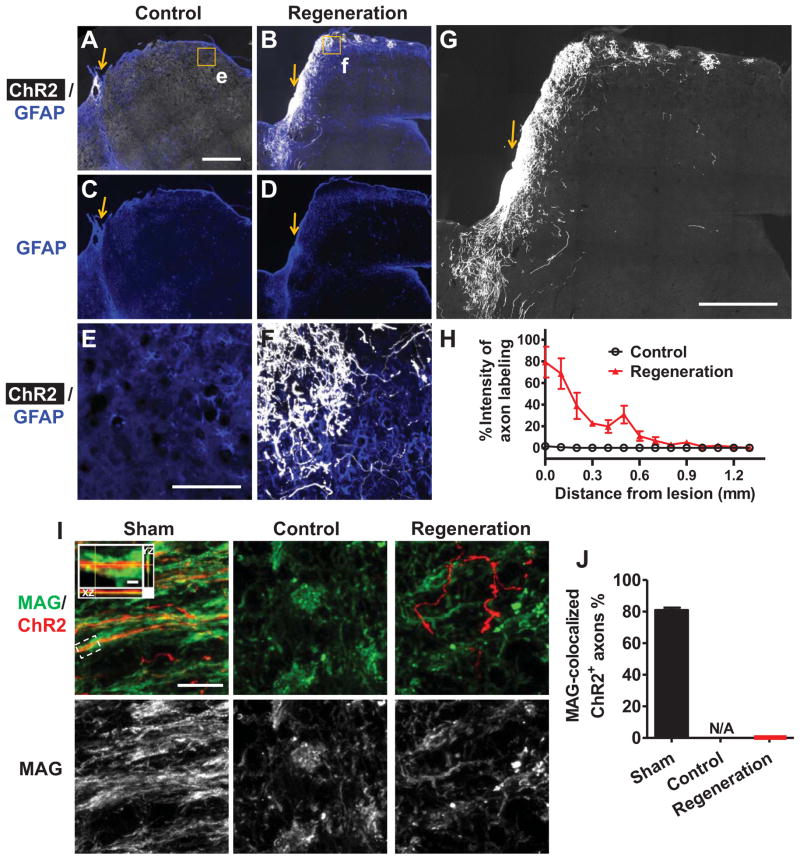
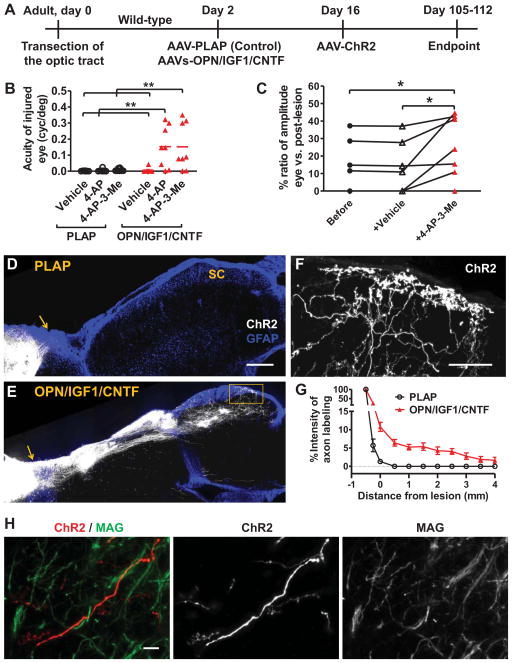
Although a number of repair strategies have been shown to promote axon outgrowth following neuronal injury in the mammalian CNS, it remains unclear whether regenerated axons establish functional synapses and support behavior. Here, in both juvenile and adult mice, we show that either PTEN and SOCS3 co-deletion, or co-overexpression of osteopontin (OPN)/insulin-like growth factor 1 (IGF1)/ciliary neurotrophic factor (CNTF), induces regrowth of retinal axons and formation of functional synapses in the superior colliculus (SC) but not significant recovery of visual function. Further analyses suggest that regenerated axons fail to conduct action potentials from the eye to the SC due to lack of myelination. Consistent with this idea, administration of voltage-gated potassium channel blockers restores conduction and results in increased visual acuity. Thus, enhancing both regeneration and conduction effectively improves function after retinal axon injury.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
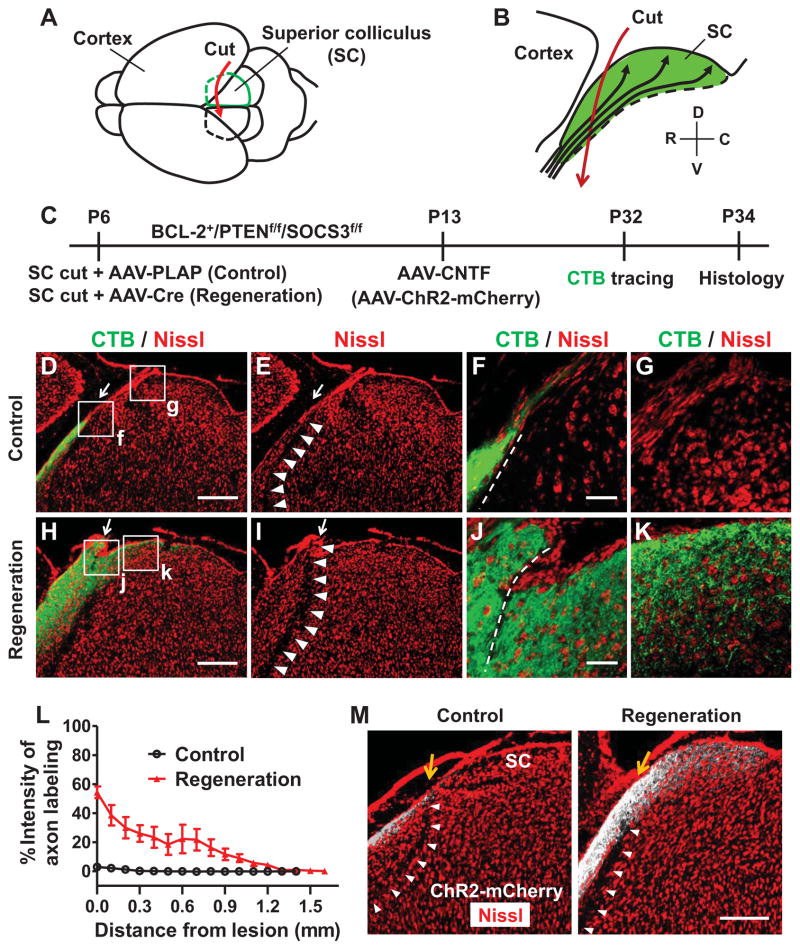
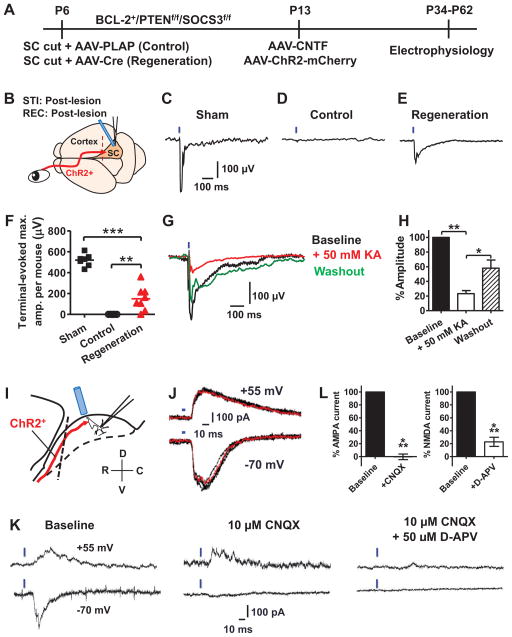
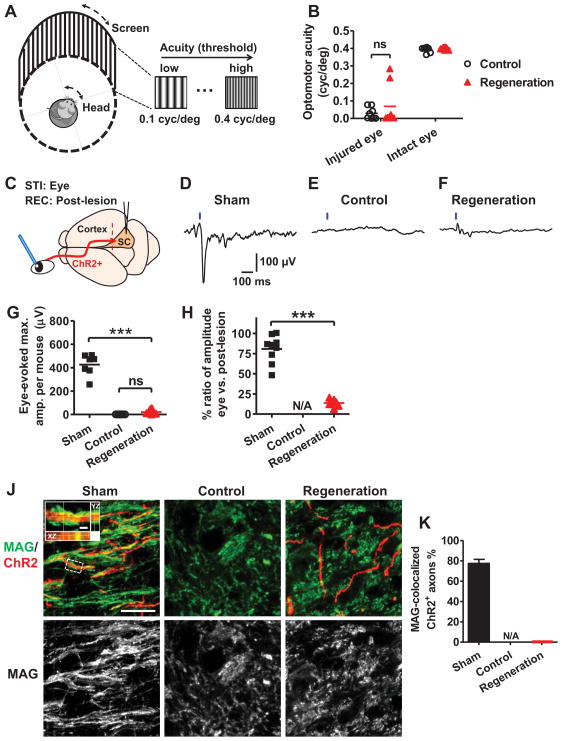
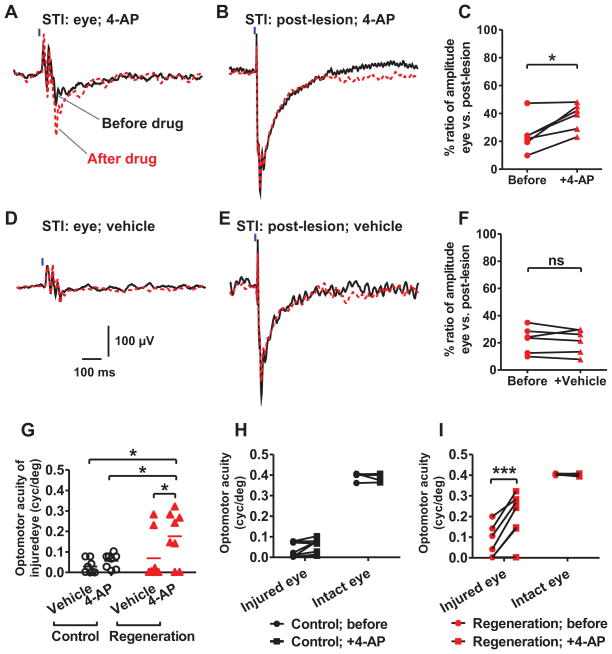
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
