

Efficient generation of Rosa26 knock-in mice using CRISPR/Cas9 in C57BL/6 zygotes
- PMID: 26772810
- PMCID: PMC4715285
- DOI: 10.1186/s12896-016-0234-4
Efficient generation of Rosa26 knock-in mice using CRISPR/Cas9 in C57BL/6 zygotes
Abstract
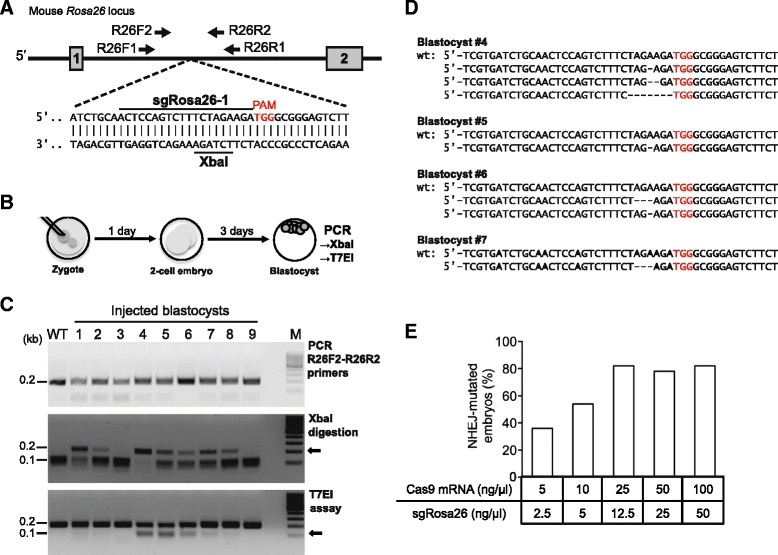
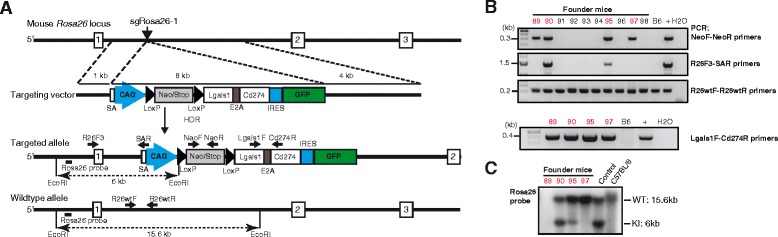
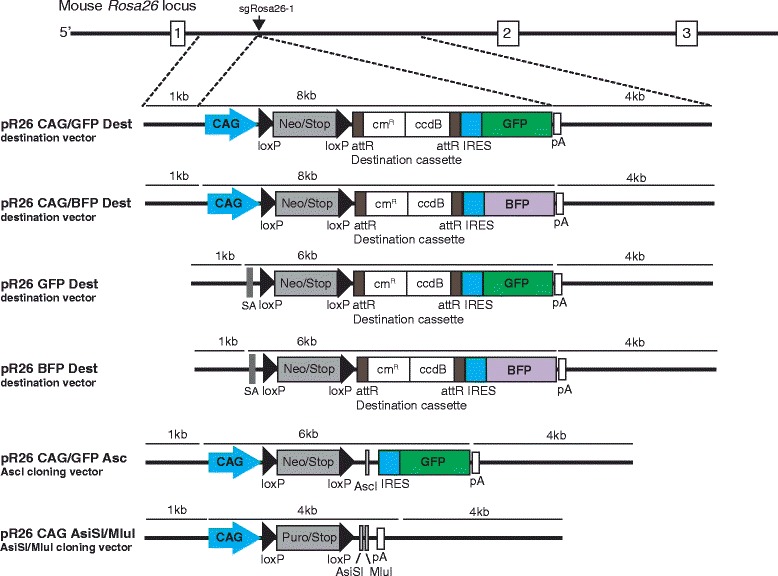
Background: The CRISPR/Cas9 system is increasingly used for gene inactivation in mouse zygotes, but homology-directed mutagenesis and use of inbred embryos are less established. In particular, Rosa26 knock-in alleles for the insertion of transgenes in a genomic 'safe harbor' site, have not been produced. Here we applied CRISPR/Cas9 for the knock-in of 8-11 kb inserts into Rosa26 of C57BL/6 zygotes.
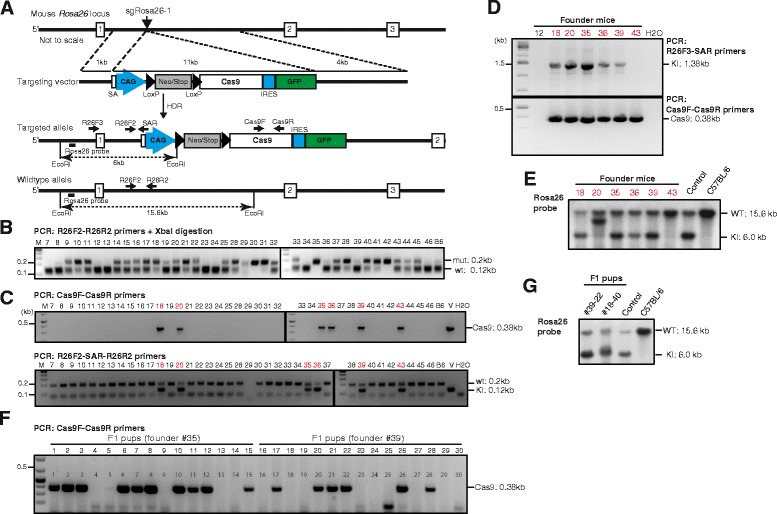
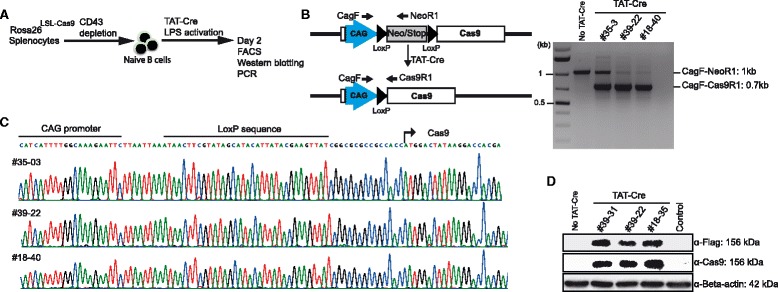
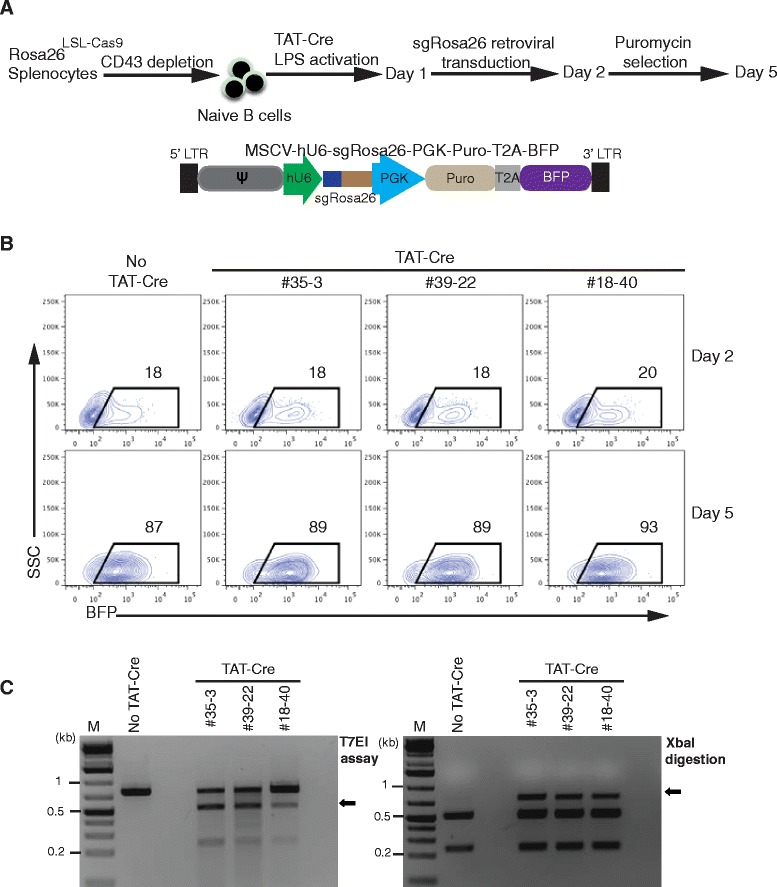
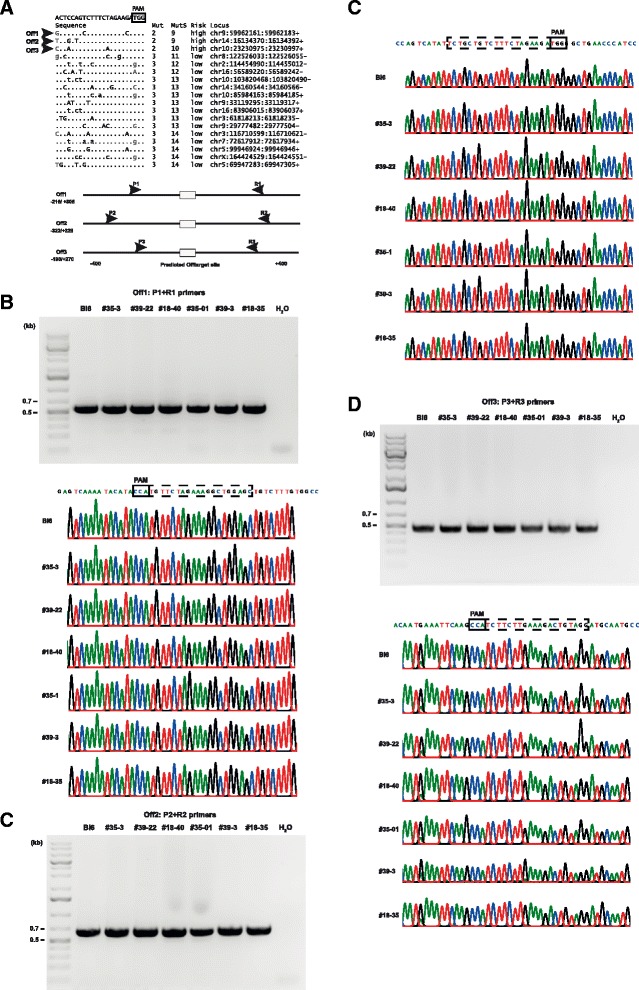
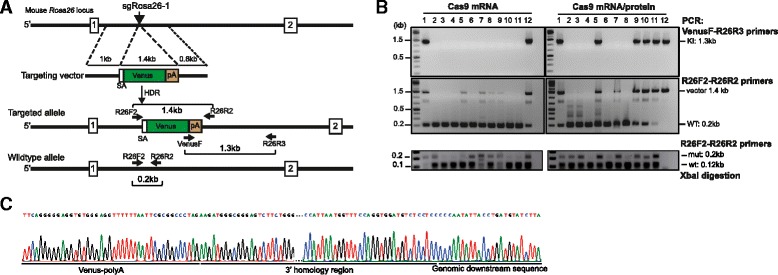
Results: We found that 10-20 % of live pups derived from microinjected zygotes were founder mutants, without apparent off-target effects, and up to 50 % knock-in embryos were recovered upon coinjection of Cas9 mRNA and protein. Using this approach, we established a new mouse line for the Cre/loxP-dependent expression of Cas9.
Conclusions: Altogether, our protocols and resources support the fast and direct generation of new Rosa26 knock-in alleles and of Cas9-mediated in vivo gene editing in the widely used C57BL/6 inbred strain.
Figures
References
-
- Zambrowicz BP, Imamoto A, Fiering S, Herzenberg LA, Kerr WG, Soriano P. Disruption of overlapping transcripts in the ROSA beta geo 26 gene trap strain leads to widespread expression of beta-galactosidase in mouse embryos and hematopoietic cells. Proc Natl Acad Sci U S A. 1997;94:3789–94. doi: 10.1073/pnas.94.8.3789. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
