

Identifying Family-Member-Specific Targets of Mono-ARTDs by Using a Chemical Genetics Approach
- PMID: 26774478
- PMCID: PMC5423403
- DOI: 10.1016/j.celrep.2015.12.045
Identifying Family-Member-Specific Targets of Mono-ARTDs by Using a Chemical Genetics Approach
Abstract
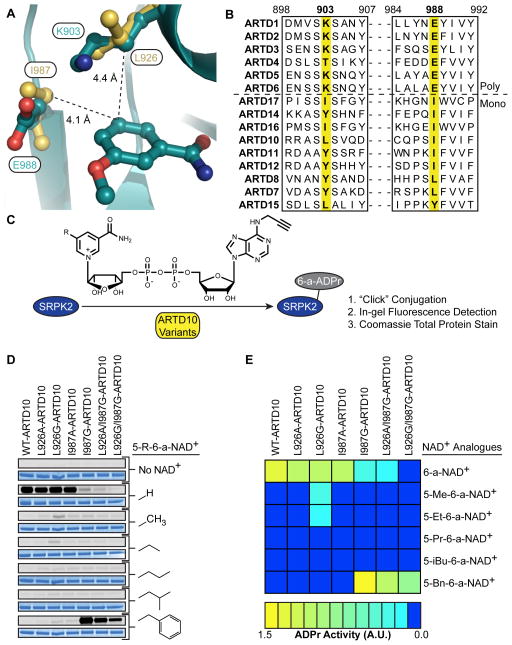
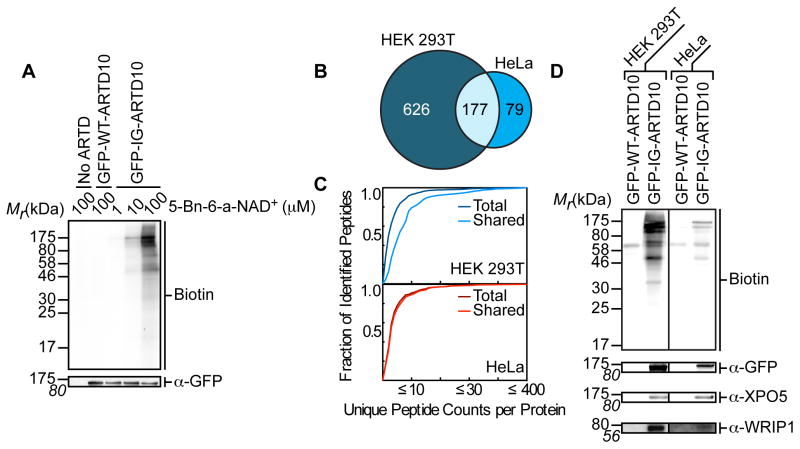
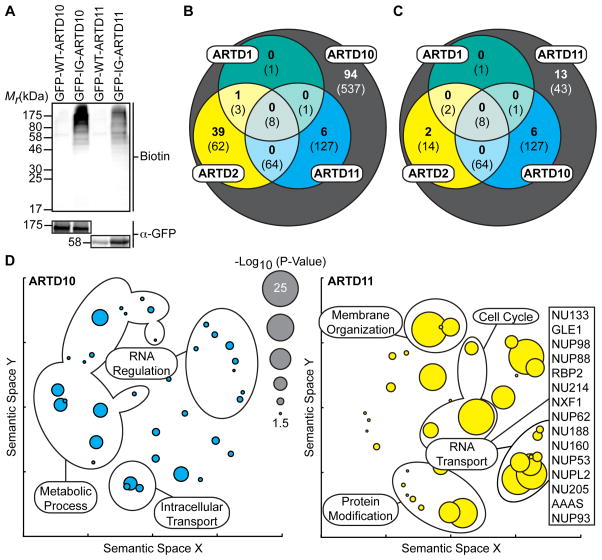
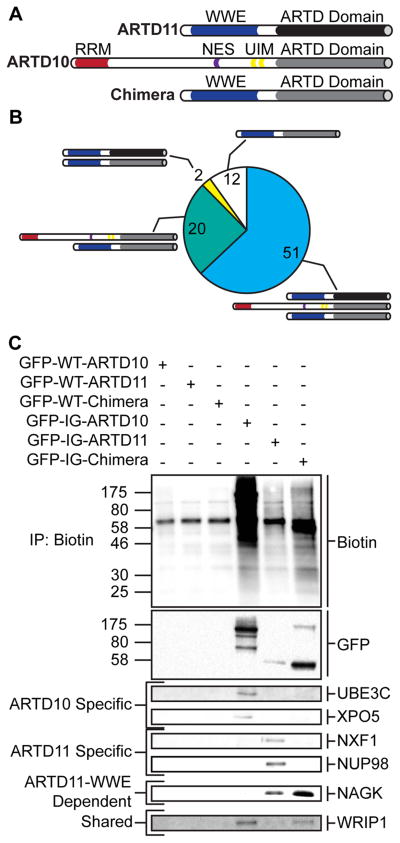
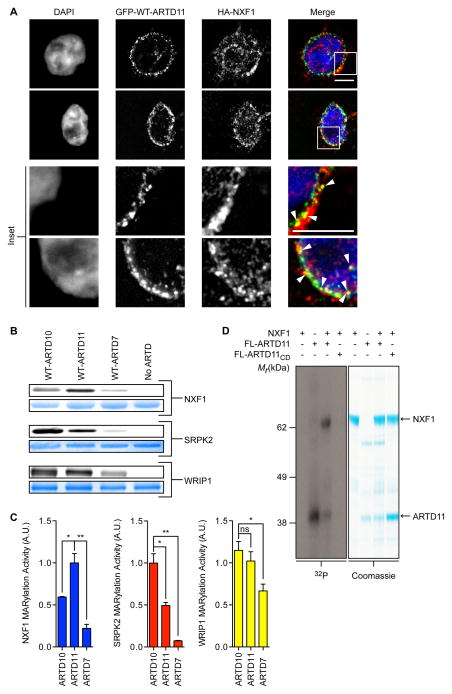
ADP-ribosyltransferases (ARTD1-16) have emerged as major downstream effectors of NAD(+) signaling in the cell. Most ARTDs (ARTD7 and 8, 10-12, and 14-17) catalyze the transfer of a single unit of ADP-ribose from NAD(+) to target proteins, a process known as mono-ADP-ribosylation (MARylation). Progress in understanding the cellular functions of MARylation has been limited by the inability to identify the direct targets for individual mono-ARTDs. Here, we engineered mono-ARTDs to use an NAD(+) analog that is orthogonal to wild-type ARTDs. We profiled the MARylomes of ARTD10 and ARTD11 in vitro, identifying isoform-specific targets and revealing a potential role for ARTD11 in nuclear pore complex biology. We found that ARTD11 targeting is dependent on both its regulatory and catalytic domains, which has important implications for how ARTDs recognize their targets. We anticipate that our chemical genetic strategy will be generalizable to all mono-ARTD family members based on the similarity of the mono-ARTD catalytic domains.
Copyright © 2016 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Ame JC, Spenlehauer C, de Murcia G. The PARP superfamily. BioEssays : news and reviews in molecular, cellular and developmental biology. 2004;26:882–893. - PubMed
-
- Cosi C, Marien M. Implication of poly (ADP-ribose) polymerase (PARP) in neurodegeneration and brain energy metabolism. Decreases in mouse brain NAD+ and ATP caused by MPTP are prevented by the PARP inhibitor benzamide. Annals of the New York Academy of Sciences. 1999;890:227–239. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
