

An optimized surgical approach for obtaining stable extracellular single-unit recordings from the cerebellum of head-fixed behaving mice
- PMID: 26777474
- PMCID: PMC4778558
- DOI: 10.1016/j.jneumeth.2016.01.010
An optimized surgical approach for obtaining stable extracellular single-unit recordings from the cerebellum of head-fixed behaving mice
Abstract
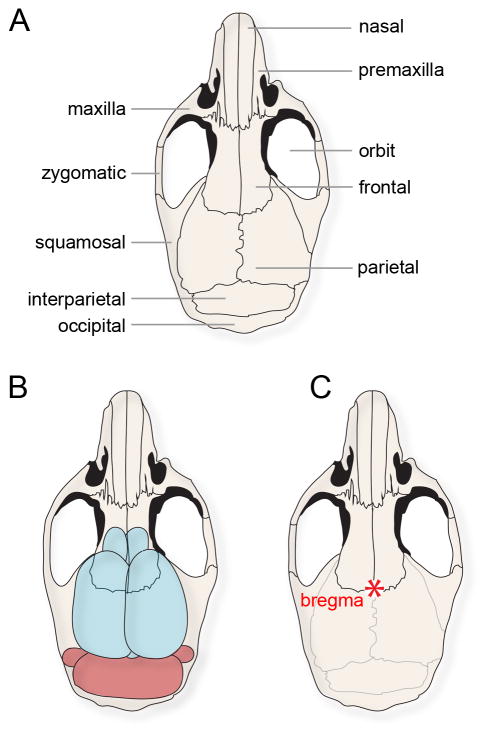
Background: Electrophysiological recording approaches are essential for understanding brain function. Among these approaches are various methods of performing single-unit recordings. However, a major hurdle to overcome when recording single units in vivo is stability. Poor stability results in a low signal-to-noise ratio, which makes it challenging to isolate neuronal signals. Proper isolation is needed for differentiating a signal from neighboring cells or the noise inherent to electrophysiology. Insufficient isolation makes it impossible to analyze full action potential waveforms. A common source of instability is an inadequate surgery. Problems during surgery cause blood loss, tissue damage and poor healing of the surrounding tissue, limited access to the target brain region, and, importantly, unreliable fixation points for holding the mouse's head.
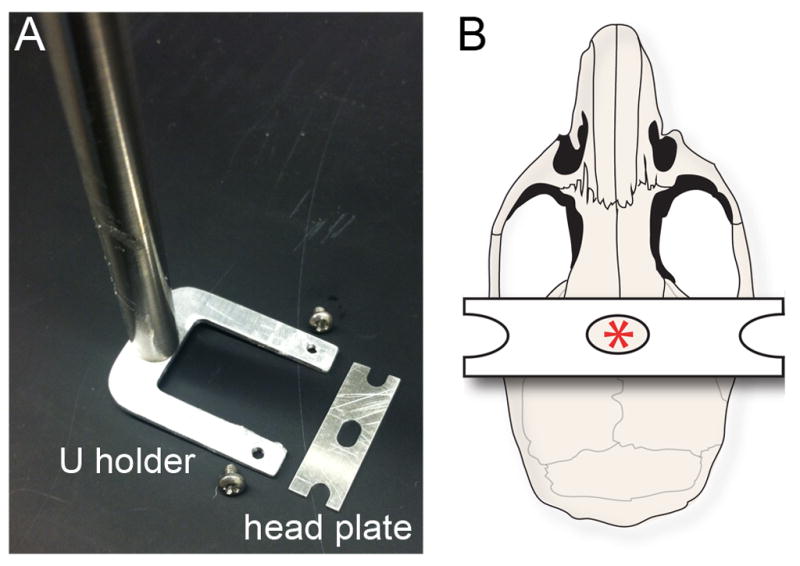
New method: We describe an optimized surgical procedure that ensures limited tissue damage and delineate a method for implanting head plates to hold the animal firmly in place.
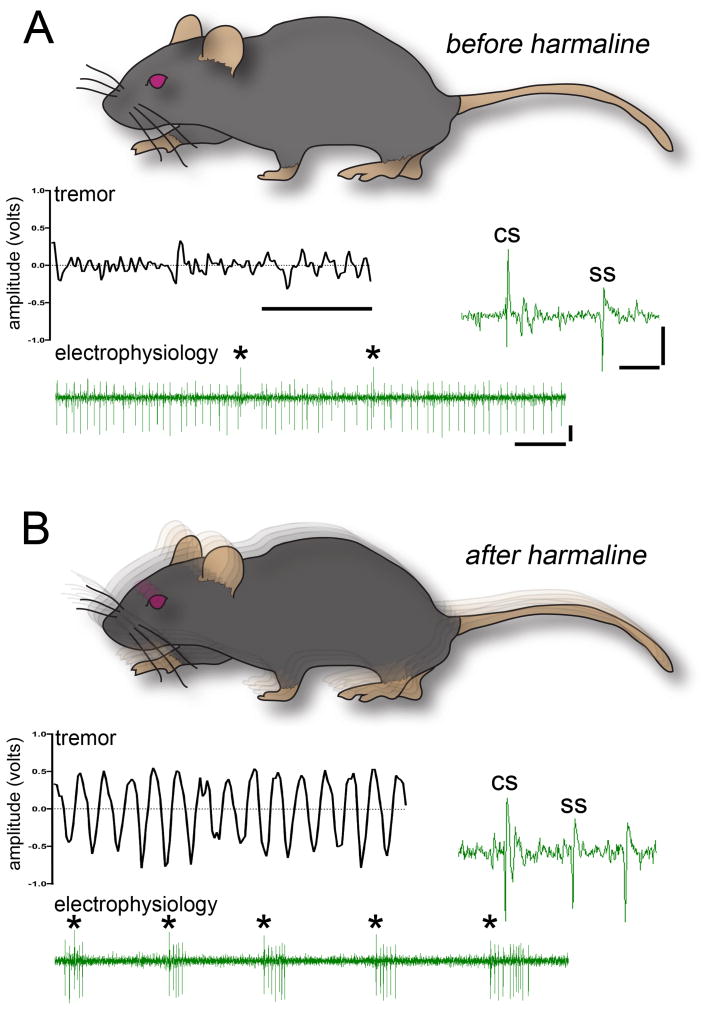
Results: Using the cerebellum as a model, we implement an extracellular recording technique to acquire single units from Purkinje cells and cerebellar nuclear neurons in behaving mice. We validate the stability of our method by holding single units after injecting the powerful tremorgenic drug harmaline. We performed multiple structural analyses after recording.
Comparison with existing methods: Our approach is ideal for studying neuronal function in active mice and valuable for recording single-neuron activity when considerable motion is unavoidable.
Conclusions: The surgical principles we present for accessing the cerebellum can be easily adapted to examine the function of neurons in other brain regions.
Keywords: Action potential; Behavior; Cerebellum; Electrophysiology; Surgery; Tremor.
Copyright © 2016 Elsevier B.V. All rights reserved.
Conflict of interest statement
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
