

A chemical proteomic atlas of brain serine hydrolases identifies cell type-specific pathways regulating neuroinflammation
- PMID: 26779719
- PMCID: PMC4737654
- DOI: 10.7554/eLife.12345
A chemical proteomic atlas of brain serine hydrolases identifies cell type-specific pathways regulating neuroinflammation
Abstract
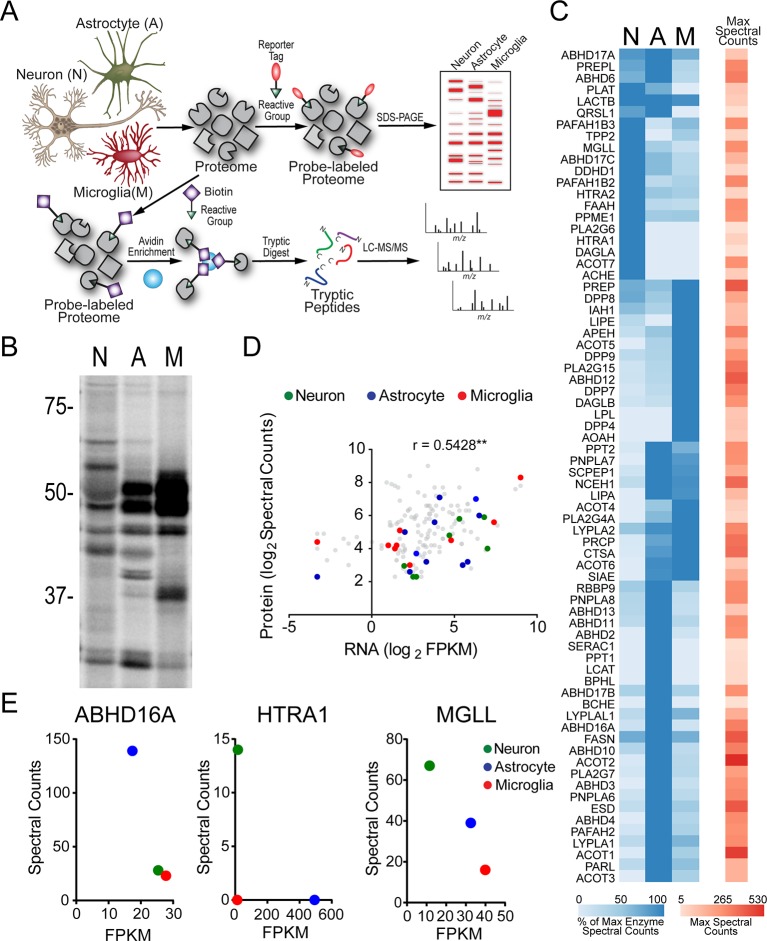
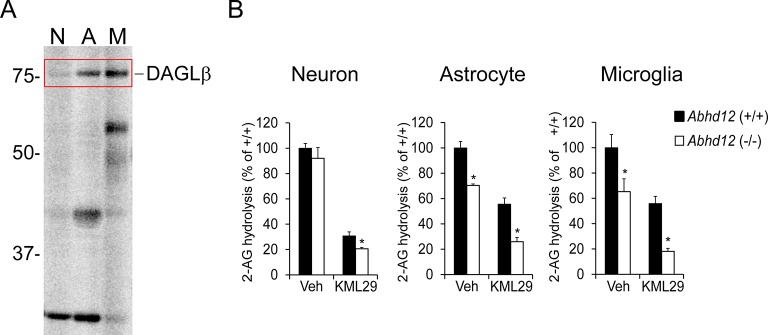
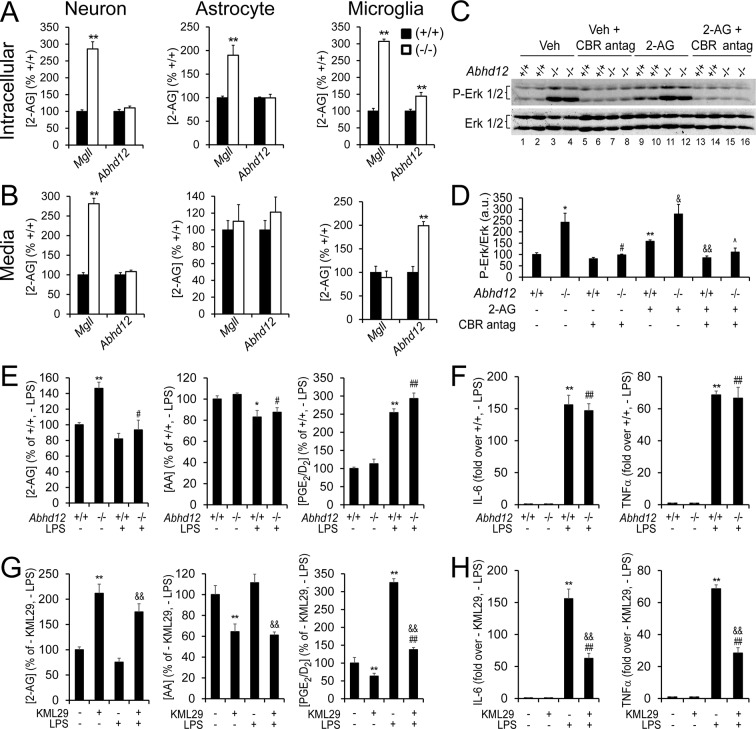
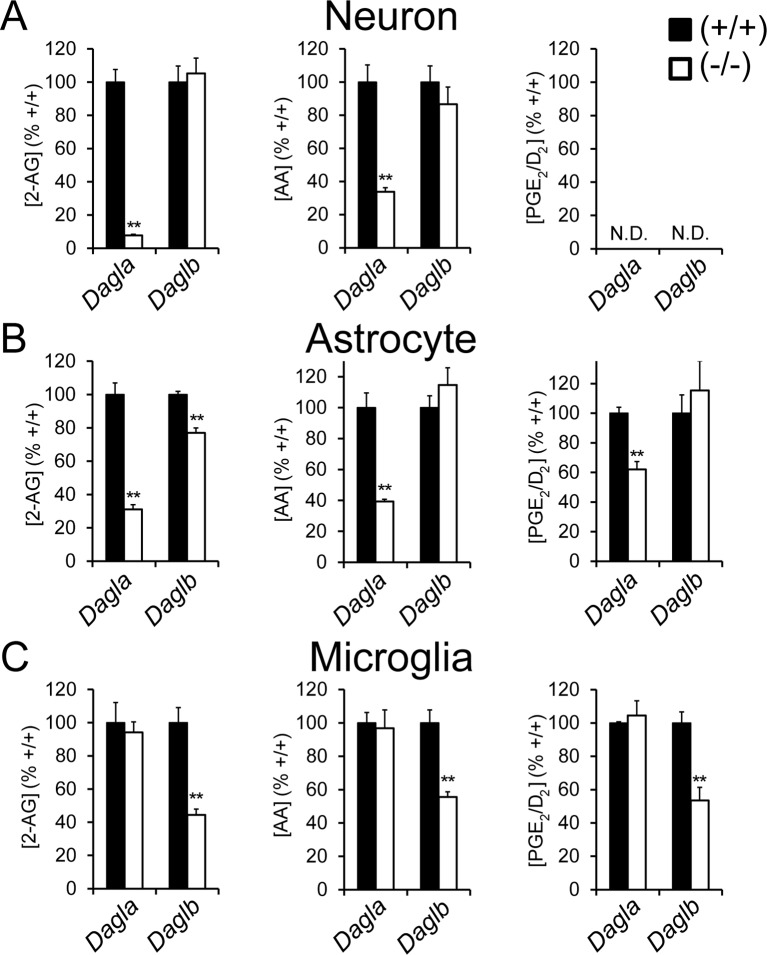
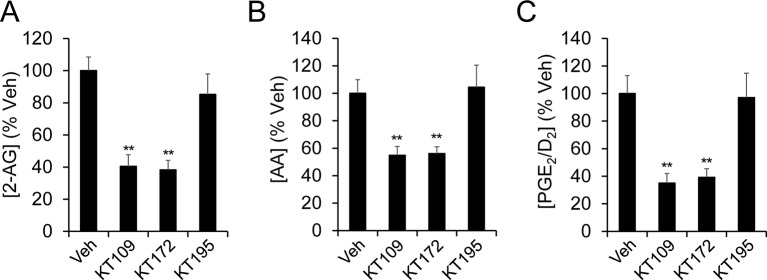
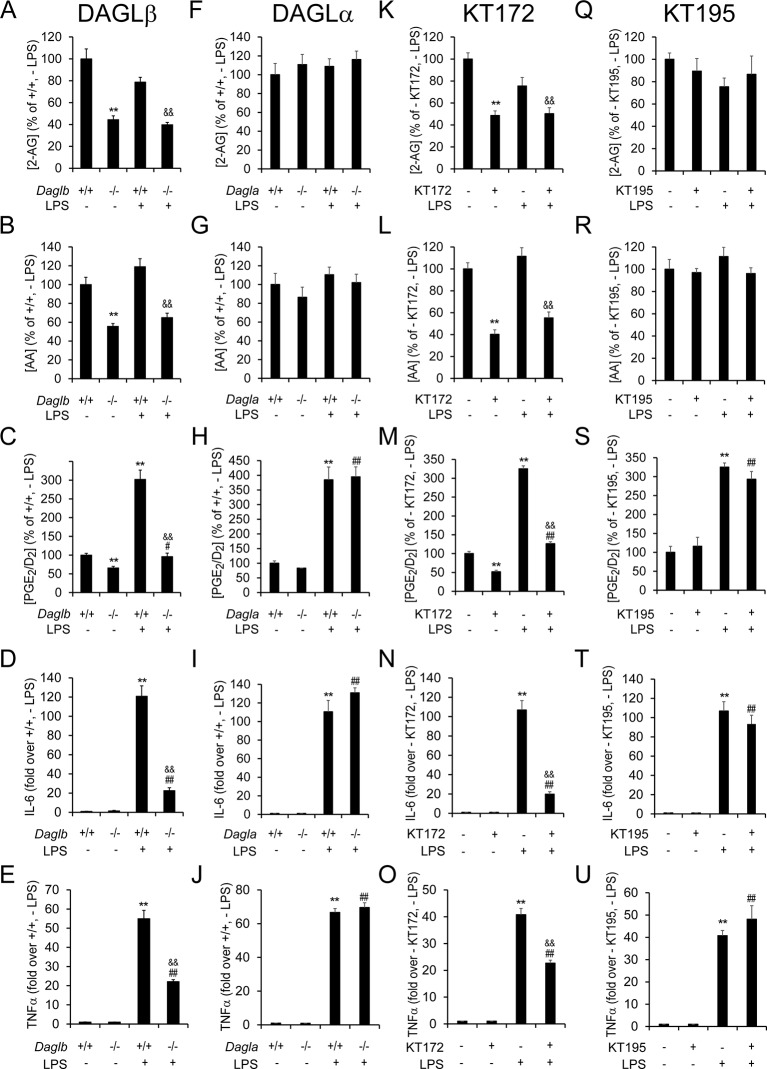
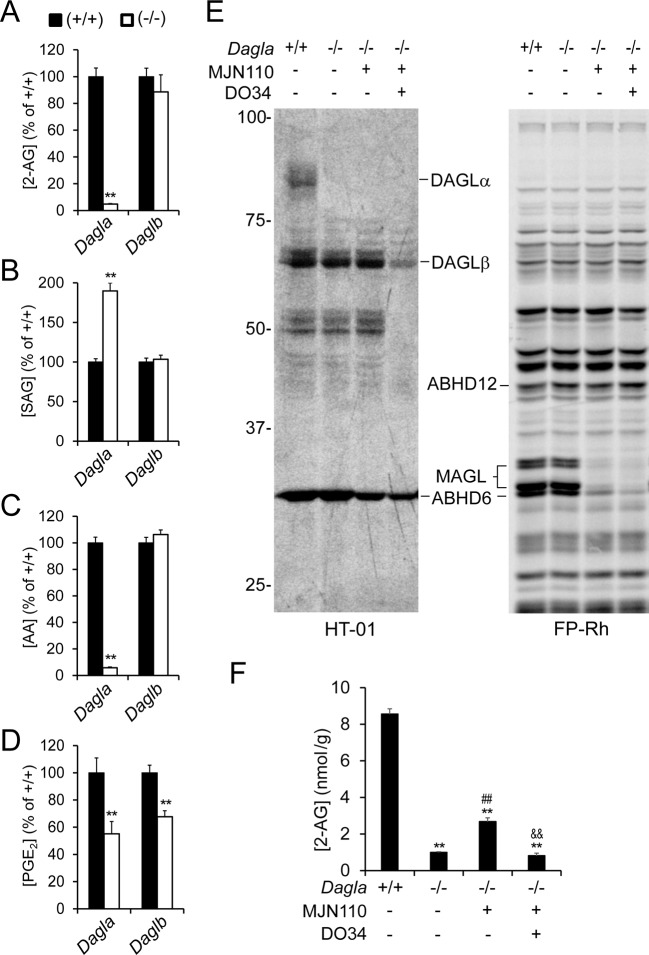
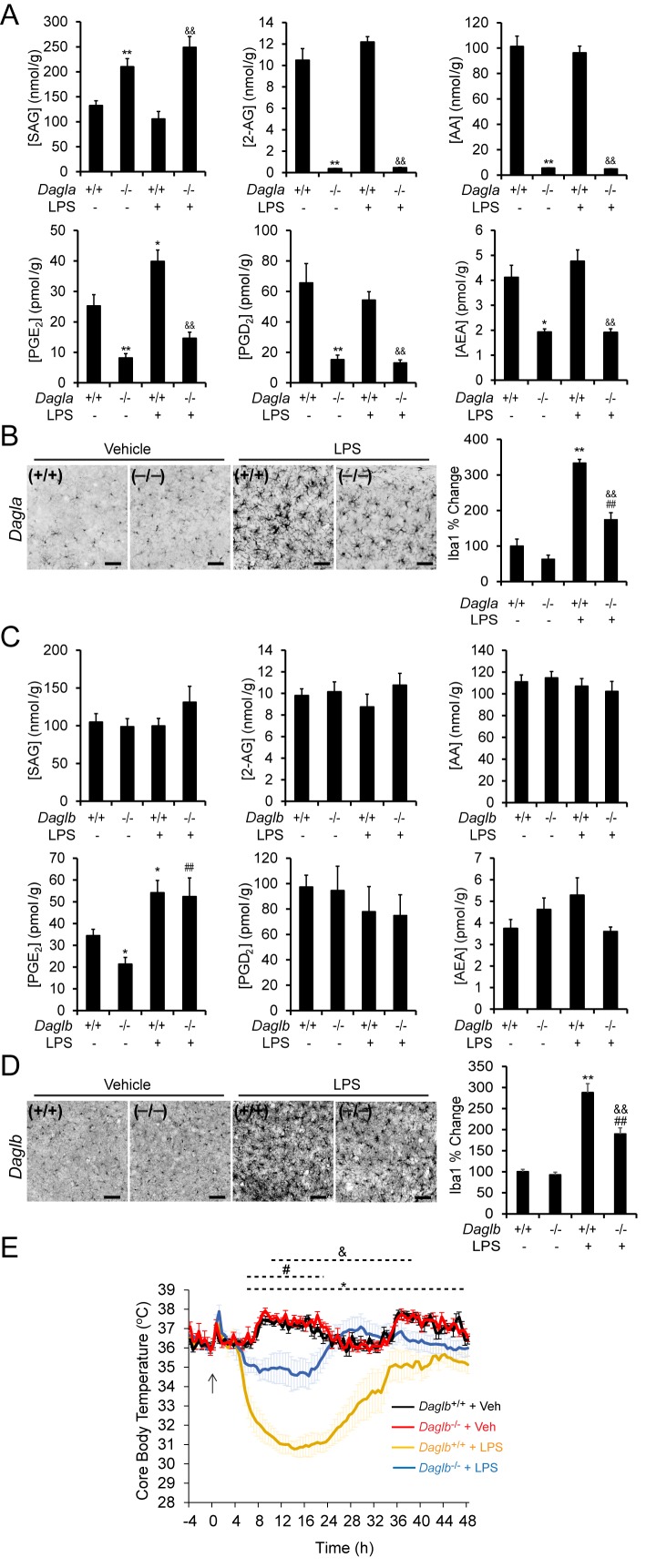
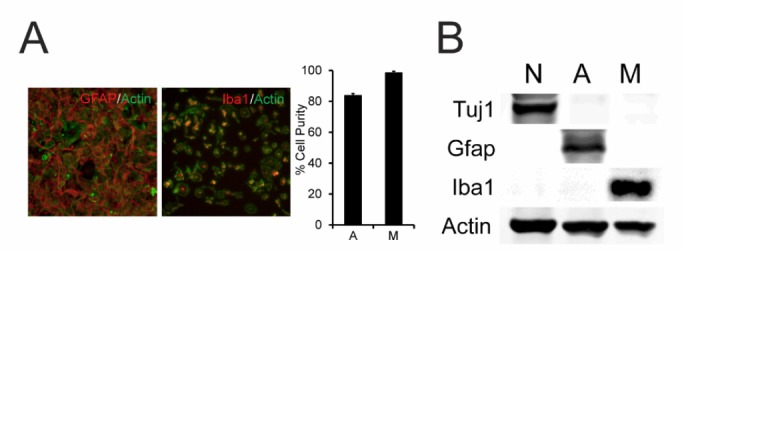
Metabolic specialization among major brain cell types is central to nervous system function and determined in large part by the cellular distribution of enzymes. Serine hydrolases are a diverse enzyme class that plays fundamental roles in CNS metabolism and signaling. Here, we perform an activity-based proteomic analysis of primary mouse neurons, astrocytes, and microglia to furnish a global portrait of the cellular anatomy of serine hydrolases in the brain. We uncover compelling evidence for the cellular compartmentalization of key chemical transmission pathways, including the functional segregation of endocannabinoid (eCB) biosynthetic enzymes diacylglycerol lipase-alpha (DAGLα) and -beta (DAGLβ) to neurons and microglia, respectively. Disruption of DAGLβ perturbed eCB-eicosanoid crosstalk specifically in microglia and suppressed neuroinflammatory events in vivo independently of broader effects on eCB content. Mapping the cellular distribution of metabolic enzymes thus identifies pathways for regulating specialized inflammatory responses in the brain while avoiding global alterations in CNS function.
Keywords: 2-arachidonoylglycerol; biochemistry; chemical proteomics; endocannabinoids; microglia; neuroinflammation; neuroscience.
Conflict of interest statement
BFC: Reviewing editor,
The other authors declares that no competing interests exist.
Figures


Similar articles
-
Inhibition of diacylglycerol lipase β modulates lipid and endocannabinoid levels in the ex vivo human placenta.Front Endocrinol (Lausanne). 2023 Feb 14;14:1092024. doi: 10.3389/fendo.2023.1092024. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 36864832 Free PMC article.
-
Rapid and profound rewiring of brain lipid signaling networks by acute diacylglycerol lipase inhibition.Proc Natl Acad Sci U S A. 2016 Jan 5;113(1):26-33. doi: 10.1073/pnas.1522364112. Epub 2015 Dec 14. Proc Natl Acad Sci U S A. 2016. PMID: 26668358 Free PMC article.
-
Endocannabinoid biosynthetic enzymes regulate pain response via LKB1-AMPK signaling.Proc Natl Acad Sci U S A. 2023 Dec 26;120(52):e2304900120. doi: 10.1073/pnas.2304900120. Epub 2023 Dec 18. Proc Natl Acad Sci U S A. 2023. PMID: 38109529 Free PMC article.
-
DAGL-dependent endocannabinoid signalling: roles in axonal pathfinding, synaptic plasticity and adult neurogenesis.Eur J Neurosci. 2011 Nov;34(10):1634-46. doi: 10.1111/j.1460-9568.2011.07831.x. Eur J Neurosci. 2011. PMID: 22103420 Review.
-
Astrocytic crosstalk with brain and immune cells in healthy and diseased conditions.Curr Opin Neurobiol. 2024 Feb;84:102840. doi: 10.1016/j.conb.2024.102840. Epub 2024 Jan 29. Curr Opin Neurobiol. 2024. PMID: 38290370 Review.
Cited by
-
Elevated Levels of Arachidonic Acid-Derived Lipids Including Prostaglandins and Endocannabinoids Are Present Throughout ABHD12 Knockout Brains: Novel Insights Into the Neurodegenerative Phenotype.Front Mol Neurosci. 2019 May 31;12:142. doi: 10.3389/fnmol.2019.00142. eCollection 2019. Front Mol Neurosci. 2019. PMID: 31213981 Free PMC article.
-
Covalent-fragment screening identifies selective inhibitors of multiple Staphylococcus aureus serine hydrolases important for growth and biofilm formation.Res Sq [Preprint]. 2024 Dec 13:rs.3.rs-5494070. doi: 10.21203/rs.3.rs-5494070/v1. Res Sq. 2024. Update in: Nat Commun. 2025 May 30;16(1):5046. doi: 10.1038/s41467-025-60367-3. PMID: 39711551 Free PMC article. Updated. Preprint.
-
Endocannabinoid dysfunction in neurological disease: neuro-ocular DAGLA-related syndrome.Brain. 2022 Oct 21;145(10):3383-3390. doi: 10.1093/brain/awac223. Brain. 2022. PMID: 35737950 Free PMC article.
-
Overabundant endocannabinoids in neurons are detrimental to cognitive function.bioRxiv [Preprint]. 2024 Sep 17:2024.09.17.613513. doi: 10.1101/2024.09.17.613513. bioRxiv. 2024. PMID: 39345517 Free PMC article. Preprint.
-
Preclinical pharmacological evaluation of the fatty acid amide hydrolase inhibitor BIA 10-2474.Br J Pharmacol. 2020 May;177(9):2123-2142. doi: 10.1111/bph.14973. Epub 2020 Feb 12. Br J Pharmacol. 2020. PMID: 31901141 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
