

Neutrophil extracellular traps enriched in oxidized mitochondrial DNA are interferogenic and contribute to lupus-like disease
- PMID: 26779811
- PMCID: PMC4742415
- DOI: 10.1038/nm.4027
Neutrophil extracellular traps enriched in oxidized mitochondrial DNA are interferogenic and contribute to lupus-like disease
Abstract
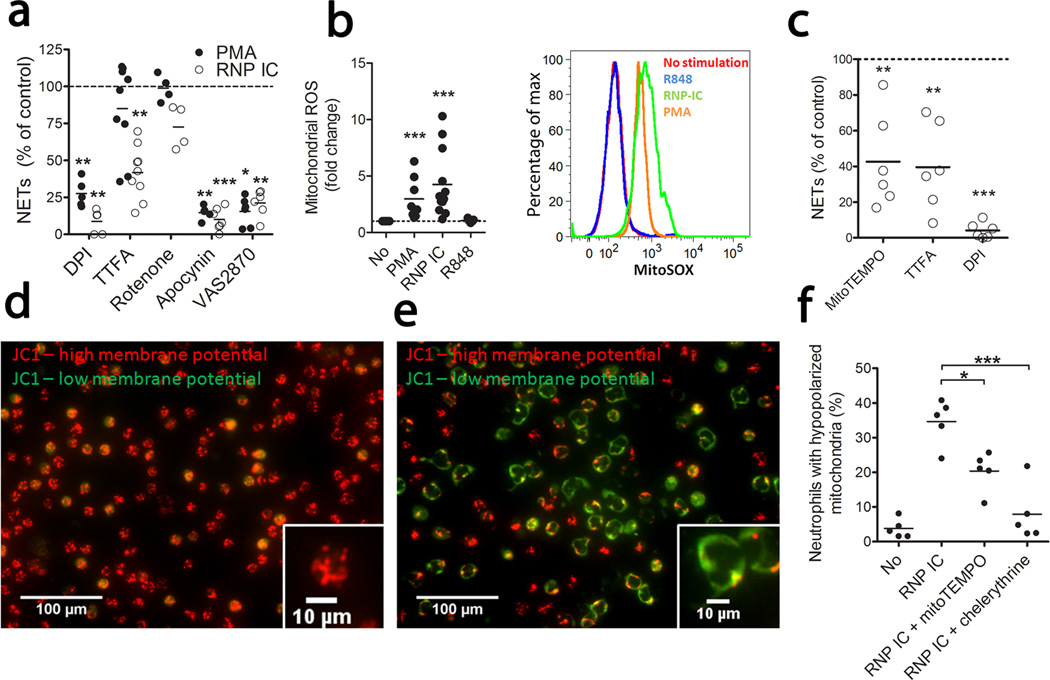
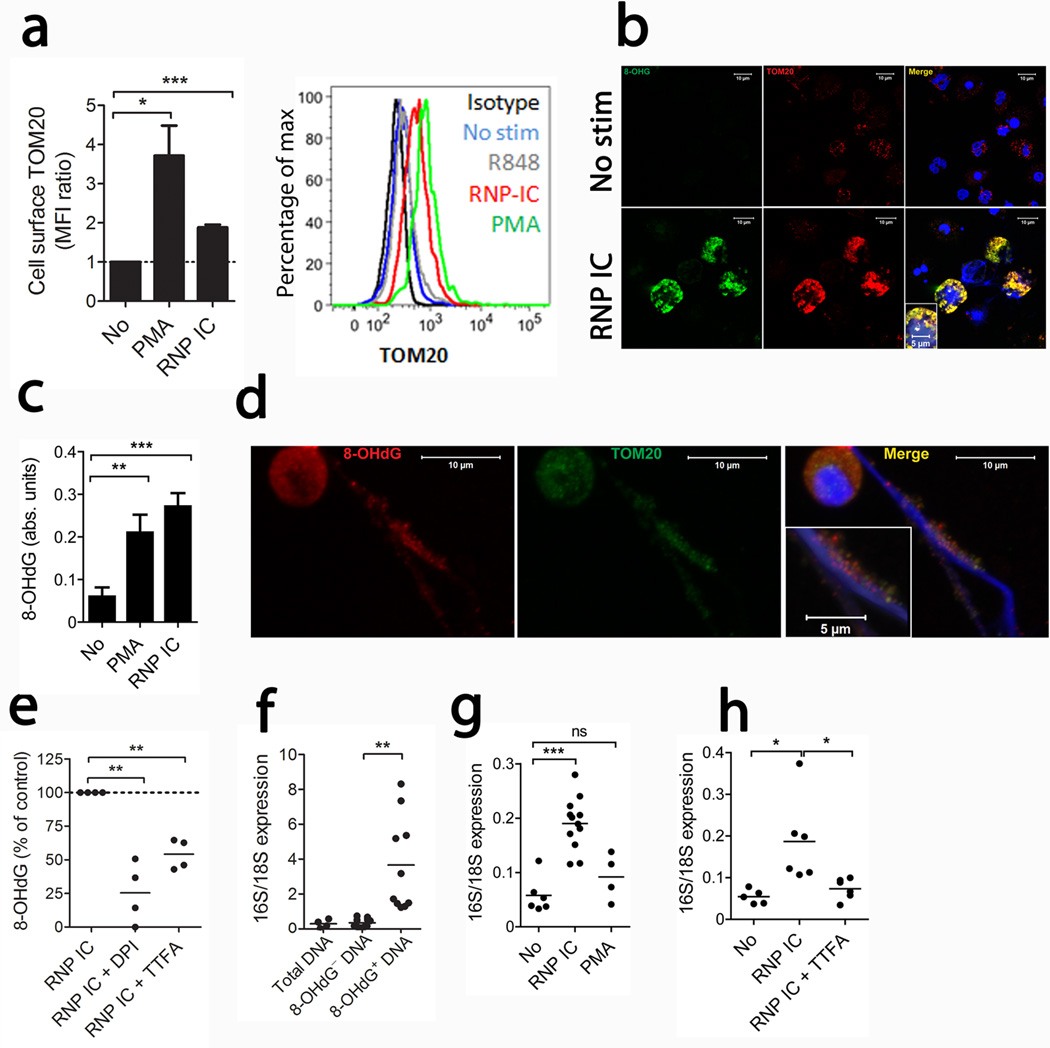
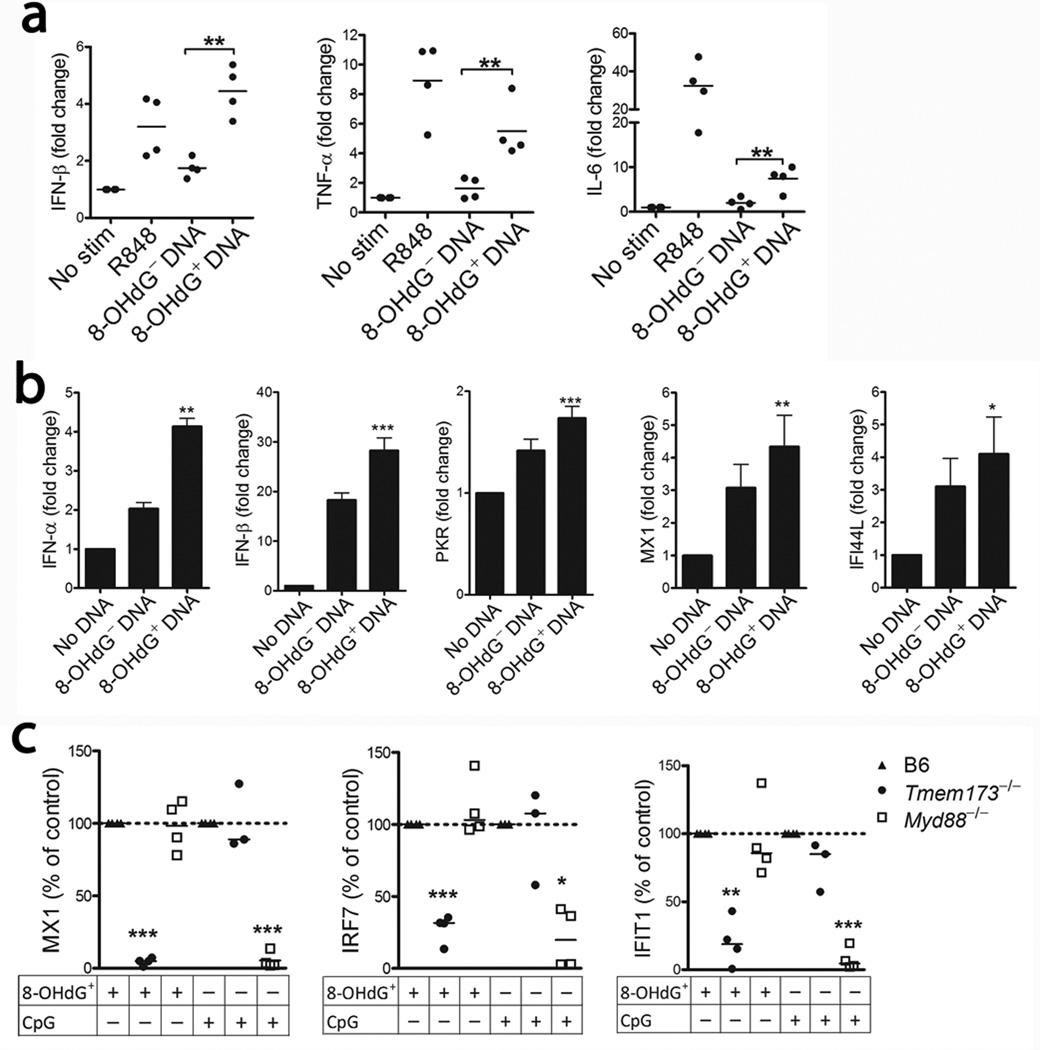
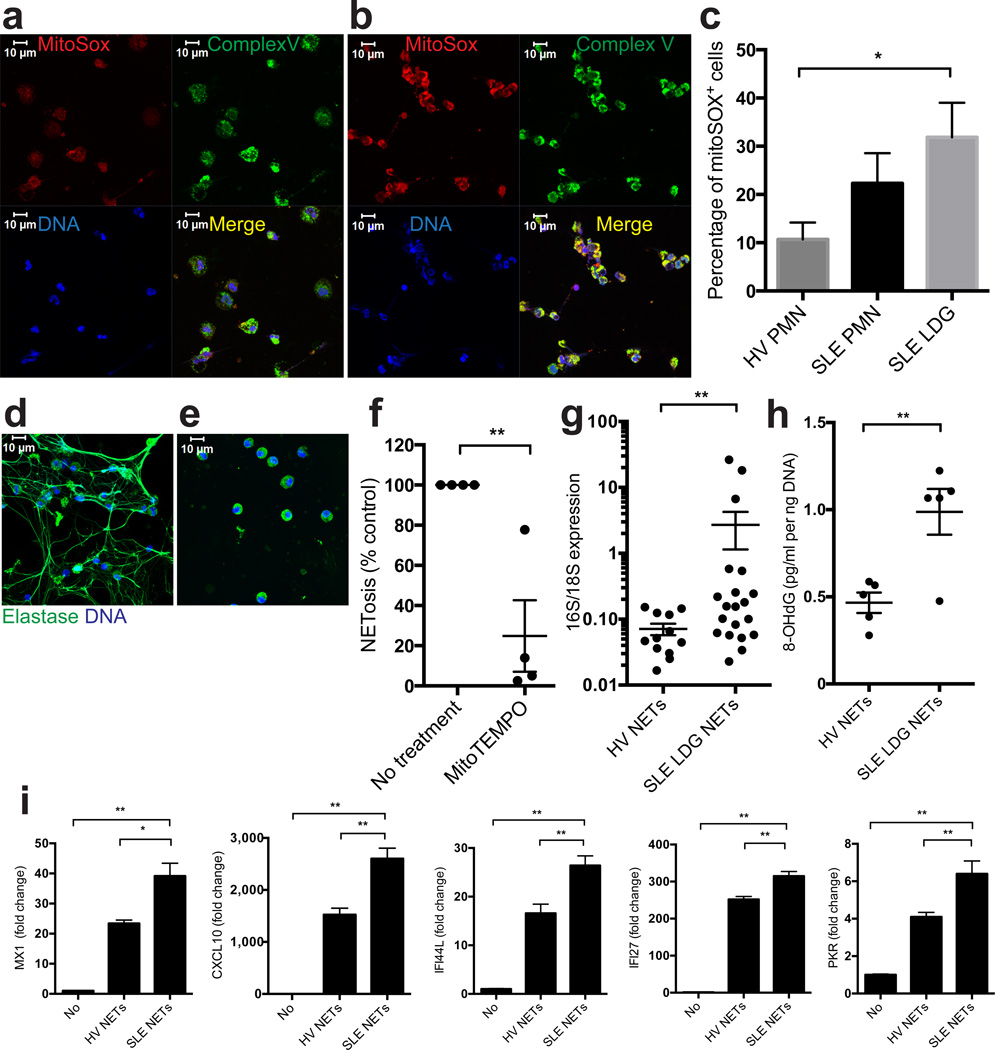
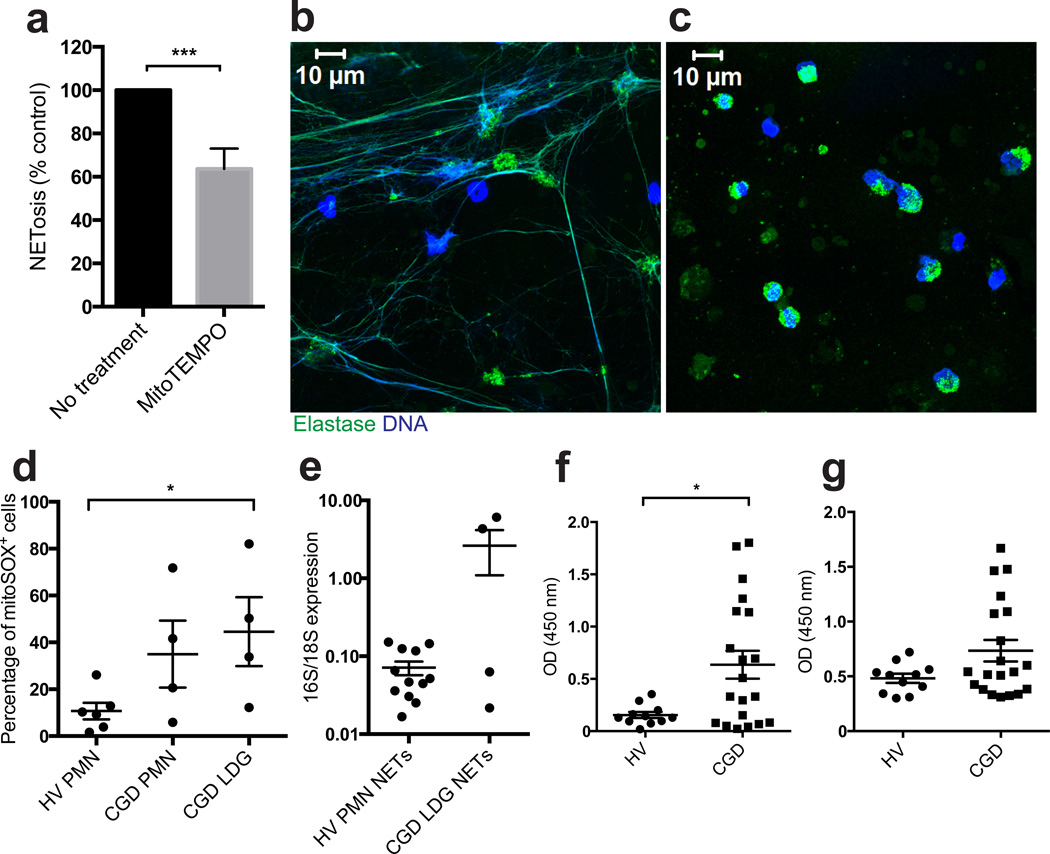
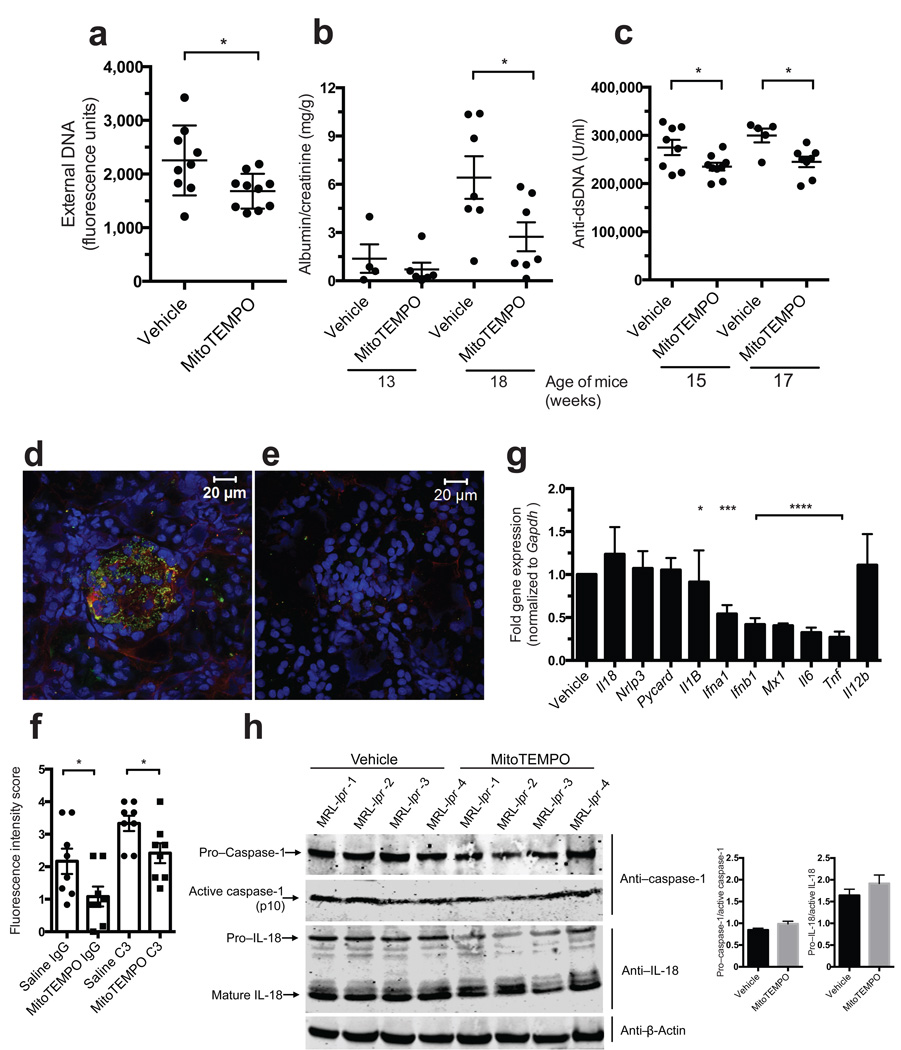
Neutrophil extracellular traps (NETs) are implicated in autoimmunity, but how they are generated and their roles in sterile inflammation remain unclear. Ribonucleoprotein immune complexes (RNP ICs), inducers of NETosis, require mitochondrial reactive oxygen species (ROS) for maximal NET stimulation. After RNP IC stimulation of neutrophils, mitochondria become hypopolarized and translocate to the cell surface. Extracellular release of oxidized mitochondrial DNA is proinflammatory in vitro, and when this DNA is injected into mice, it stimulates type I interferon (IFN) signaling through a pathway dependent on the DNA sensor STING. Mitochondrial ROS are also necessary for spontaneous NETosis of low-density granulocytes from individuals with systemic lupus erythematosus. This was also observed in individuals with chronic granulomatous disease, who lack NADPH oxidase activity but still develop autoimmunity and type I IFN signatures. Mitochondrial ROS inhibition in vivo reduces disease severity and type I IFN responses in a mouse model of lupus. Together, these findings highlight a role for mitochondria in the generation not only of NETs but also of pro-inflammatory oxidized mitochondrial DNA in autoimmune diseases.
Conflict of interest statement
The authors have declared that no conflicts of interest exist.
Figures
Comment in
-
Oxidation and mitochondrial origin of NET DNA in the pathogenesis of lupus.Nat Med. 2016 Feb;22(2):126-7. doi: 10.1038/nm.4044. Nat Med. 2016. PMID: 26845404 No abstract available.
-
Connective tissue diseases: Mitochondria drive NETosis and inflammation in SLE.Nat Rev Rheumatol. 2016 Apr;12(4):195-6. doi: 10.1038/nrrheum.2016.24. Epub 2016 Mar 3. Nat Rev Rheumatol. 2016. PMID: 26935279
References
-
- Nathan C. Neutrophils and immunity: challenges and opportunities. Nat Rev Immunol. 2006;6:173–182. - PubMed
-
- Kolaczkowska E, Kubes P. Neutrophil recruitment and function in health and inflammation. Nat Rev Immunol. 2013;13:159–175. - PubMed
-
- Brinkmann V, et al. Neutrophil extracellular traps kill bacteria. Science. 2004;303:1532–1535. - PubMed
Methods-only references
-
- Tan EM, et al. The 1982 revised criteria for the classification of systemic lupus erythematosus. Arthritis Rheum. 1982;25:1271–1277. - PubMed
-
- Olferiev M, Lliguicota M, Kirou KA, Crow MK. Measuring interferon alpha and other cytokines in SLE. Methods Mol Biol. 2014;1134:131–150. - PubMed
-
- Ahmad S, Ghosh A, Nair DL, Seshadri M. Simultaneous extraction of nuclear and mitochondrial DNA from human blood. Genes Genet Syst. 2007;82:429–432. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
